Onda beta
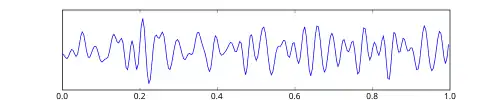
As ondas beta, ou ritmo beta, são oscilações neurais (ondas cerebrais) no cérebro humano com uma faixa de frequência entre 12,5 e 30 Hz (12,5 a 30 ciclos por segundo). Vários ritmos diferentes coexistem, com alguns sendo inibitórios e outros excitatórios em função.[1]
As ondas beta podem ser divididas em três seções: Ondas Beta Baixas (12,5–16 Hz, "Beta 1"); Ondas Beta (16,5–20 Hz, "Beta 2"); e Ondas Beta Altas (20,5–28 Hz, "Beta 3").[2] Os estados beta são os estados associados à consciência de vigília normal.
História
As ondas beta foram descobertas e nomeadas pelo psiquiatra alemão Hans Berger, que inventou a eletroencefalografia (EEG) em 1924, como um método de registrar a atividade elétrica do cérebro a partir do couro cabeludo humano. Berger denominou as ondas de maior amplitude e menor frequência que apareciam na parte posterior do couro cabeludo quando os olhos do sujeito estavam fechados de ondas alfa. As ondas de menor amplitude e maior frequência que substituíam as ondas alfa quando o sujeito abria os olhos foram então denominadas ondas beta.[3]
Função
Ondas beta de baixa amplitude com frequências múltiplas e variadas são frequentemente associadas a pensamentos ativos, ocupados ou ansiosos e à concentração ativa.[4]
Sobre o córtex motor, as ondas beta estão associadas às contrações musculares que ocorrem em movimentos isotônicos [en] e são suprimidas antes e durante as mudanças de movimento,[5] com observações semelhantes em habilidades motoras finas e grossas.[6] Surtos de atividade beta estão associados a um fortalecimento do feedback sensorial no controle motor estático e são reduzidos quando há mudança de movimento.[7] A atividade beta aumenta quando o movimento precisa ser resistido ou suprimido voluntariamente.[8] A indução artificial de aumento das ondas beta sobre o córtex motor por uma forma de estimulação elétrica chamada estimulação transcraniana por corrente alternada [en], consistente com sua ligação à contração isotônica, produz uma desaceleração dos movimentos motores.[9]
Investigações sobre o feedback de recompensa revelaram dois componentes beta distintos; um componente beta alto (gama baixo),[10] e um componente beta baixo.[11] Em associação com ganhos inesperados, o componente beta alto é mais profundo ao receber um resultado inesperado, com baixa probabilidade.[12] No entanto, diz-se que o componente beta baixo está relacionado à omissão de ganhos, quando os ganhos são esperados.[11]
Durante o repouso, as oscilações beta rápidas são prevalentes sobre o córtex pré-frontal lateral [en] (CPFL) em humanos, seguindo um aumento de frequência posteroanterior. Este aumento da frequência beta em sub-regiões mais anteriores corresponde a um modelo anatômico da função do CPFL, no qual o controle cognitivo é organizado hierarquicamente, com mecanismos de controle mais abstratos e sofisticados servidos pelas regiões mais anteriores e um controle mais direto/concreto da ação direcionada a objetivos em locais posteriores. Isso está ainda de acordo com o fato de que o beta do CPFL posterior é mais lento em frequência, semelhante ao observado sobre o córtex motor em repouso.[13]
Relação com o GABA
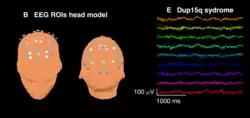
As ondas beta são frequentemente consideradas indicativas de transmissão cortical inibitória mediada pelo ácido gama-aminobutírico (GABA), o principal neurotransmissor inibitório do sistema nervoso dos mamíferos. Os benzodiazepínicos, drogas que modulam os receptores GABAA, induzem ondas beta em registros de EEG de humanos[14] e ratos.[15] Ondas beta espontâneas também são observadas difusamente em registros de EEG de couro cabeludo de crianças com síndrome de duplicação 15q11.2-q13.1 (Dup15q [en]), que têm duplicações dos genes da subunidade do receptor GABAA GABRA5 [en], GABRB3 [en] e GABRG3 [en].[16] Da mesma forma, crianças com síndrome de Angelman com deleções dos mesmos genes da subunidade do receptor GABAA apresentam amplitude beta diminuída.[17] Assim, as ondas beta são provavelmente biomarcadores de disfunção GABAérgica, especialmente em distúrbios do neurodesenvolvimento causados por deleções/duplicações 15q.
Ver também
Ondas cerebrais
- Onda delta – (0,1 – 3 Hz)
- Onda theta [en] – (4 – 7 Hz)
- Onda alfa – (7 – 12 Hz)
- Onda gama – (30 – 100 Hz)
Padre Marcelo Rossi | |||||
|---|---|---|---|---|---|
| Álbuns |
| ||||
| Publicações |
| ||||
| Filmografia |
| ||||
| Televisão |
| ||||
| Artigos relacionados |
| ||||
- ↑ Rassi, Elie; Lin, Wy Ming; Zhang, Yi; Emmerzaal, Jill; Haegens, Saskia (2023). «β Band Rhythms Influence Reaction Times». eNeuro. 10 (6). ISSN 2373-2822. PMC 10312120
 . PMID 37364994. doi:10.1523/ENEURO.0473-22.2023
. PMID 37364994. doi:10.1523/ENEURO.0473-22.2023
- ↑ Rangaswamy M, Porjesz B, Chorlian DB, Wang K, Jones KA, Bauer LO, Rohrbaugh J, O'Connor SJ, Kuperman S, Reich T, Begleiter (2002). «Beta power in the EEG of alcoholics». Biological Psychology. 52 (8): 831–842. PMID 12372655. doi:10.1016/s0006-3223(02)01362-8
- ↑ Buzsáki, György (2006). Rhythms of the Brain. Nova Iorque: Oxford University Press. p. 4. ISBN 978-0-19-530106-9
- ↑ Baumeister J, Barthel T, Geiss KR, Weiss M (2008). «Influence of phosphatidylserine on cognitive performance and cortical activity after induced stress». Nutritional Neuroscience. 11 (3): 103–110. PMID 18616866. doi:10.1179/147683008X301478
- ↑ Baker, SN (2007). «Oscillatory interactions between sensorimotor cortex and the periphery». Current Opinion in Neurobiology. 17 (6): 649–55. PMC 2428102. PMID 18339546. doi:10.1016/j.conb.2008.01.007
- ↑ Easthope, Eric; Shamei, Arian; Liu, Yadong; Gick, Bryan; Fels, Sidney (17 de agosto de 2023). «Cortical control of posture in fine motor skills: evidence from inter-utterance rest position». Frontiers in Human Neuroscience. 17. PMC 10469778. PMID 37662639. doi:10.3389/fnhum.2023.1139569
- ↑ Lalo, E; Gilbertson, T; Doyle, L; Di Lazzaro, V; Cioni, B; Brown, P (2007). «Phasic increases in cortical beta activity are associated with alterations in sensory processing in the human». Experimental Brain Research. Experimentelle Hirnforschung. Experimentation Cerebrale. 177 (1): 137–45. PMID 16972074. doi:10.1007/s00221-006-0655-8
- ↑ Zhang, Y; Chen, Y; Bressler, SL; Ding, M (2008). «Response preparation and inhibition: the role of the cortical sensorimotor beta rhythm». Neuroscience. 156 (1): 238–46. PMC 2684699. PMID 18674598. doi:10.1016/j.neuroscience.2008.06.061
- ↑ Pogosyan, A; Gaynor, LD; Eusebio, A; Brown, P (2009). «Boosting cortical activity at Beta-band frequencies slows movement in humans». Current Biology. 19 (19): 1637–41. Bibcode:2009CBio...19.1637P. PMC 2791174. PMID 19800236. doi:10.1016/j.cub.2009.07.074
- ↑ Marco-Pallerés, J., Cucurell, D., Cunillera, T., García, R., Andrés-Pueyo, A., Münte, T. F., et al. (2008).Human oscillatory activity associated to reward processing in a gambling task, Neuropsychologia, 46, 241-248. doi:10.1016/j.neuropsychologia.2007.07.016
- ↑ a b Yaple, Z., Martinez-Saito, M., Novikov, N., Altukhov, D., Shestakova, A., Klucharev, V. (2018). Power of feedback-induced beta oscillations reflect omission of rewards: Evidence from an EEG gambling study, Frontiers in Neuroscience, 12, 776. doi:10.3389/fnins.2018.00776
- ↑ HajiHosseini, A., Rodriguez-Fornells, A., and Marco-Pallerés, J. (2012). The role of beta-gamma oscillations in unexpected rewards processing, Neuroimage, 60, 1678-1685. doi:10.1016/j.neuroimage.2012.01.125
- ↑ Capilla, A., Arana, L., García-Huéscar, M., Melcón, M., Gross, J., & Campo, P. (2022). The natural frequencies of the resting human brain: An MEG-based atlas. NeuroImage, 258, 119373.
- ↑ Feshchenko, V; Veselis, R; Reinsel, R (1997). «Comparison of the EEG effects of midazolam, thiopental, and propofol: the role of underlying oscillatory systems». Neuropsychobiology. 35 (4): 211–20. PMID 9246224. doi:10.1159/000119347
- ↑ Van Lier, Hester; Drinkenburg, Wilhelmus; Van Eeten, Yvonne; Coenen, Anton (2004). «Effects of diazepam and zolpidem on EEG beta frequencies are behavior-specific in rats». Neuropharmacology. 47 (2): 163–174. PMID 15223295. doi:10.1016/j.neuropharm.2004.03.017
- ↑ Frohlich, Joel; Senturk, Damla; Saravanapandian, Vidya; Golshani, Peyman; Reiter, Lawrence; Sankar, Raman; Thibert, Ronald; DiStefano, Charlotte; Huberty, Scott; Cook, Edwin; Jeste, Shafali (dezembro de 2016). «A Quantitative Electrophysiological Biomarker of Duplication 15q11.2-q13.1 Syndrome». PLOS ONE. 11 (12). Bibcode:2016PLoSO..1167179F. PMC 5157977. PMID 27977700. doi:10.1371/journal.pone.0167179
- ↑ Hipp, Joerg F.; Khwaja, Omar; Krishnan, Michelle; Jeste, Shafali S.; Rotenberg, Alexander; Hernandez, Maria-Clemencia; Tan, Wen-Hann; Sidorov, Michael S.; Philpot, Benjamin D. (18 de janeiro de 2019). «Electrophysiological Phenotype in Angelman Syndrome Differs Between Genotypes». Biological Psychiatry (em inglês). 85 (9): 752–759. ISSN 0006-3223. PMC 6482952. PMID 30826071. doi:10.1016/j.biopsych.2019.01.008