Xenoturbella bocki
Xenoturbella bocki
| |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
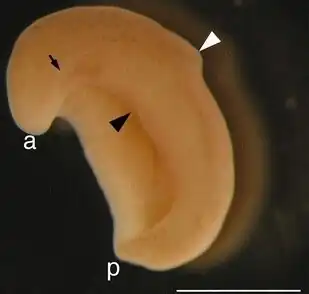 X. bocki. A seta preta indica o sulco lateral. a é a ponta anterior. p é a ponta posterior. O triângulo preto indica a boca. O triângulo branco indica o sulco circunferencial. A barra de escala no canto inferior direito mede 1 cm. | |||||||||||
| Classificação científica | |||||||||||
| |||||||||||
| Nome binomial | |||||||||||
| Xenoturbella bocki Westblad, 1949 | |||||||||||
Xenoturbella bocki é uma espécie marinha bentônica semelhante a um verme do gênero Xenoturbella. Habita ambientes de fundo oceânico em águas salgadas, principalmente ao largo da costa da Suécia, na Europa. Foi a primeira espécie do gênero descoberta, coletada inicialmente pelo zoólogo sueco Sixten Bock em 1915 e descrita em 1949 pelo zoólogo sueco Einar Westblad.[1] Sua estrutura digestiva peculiar, na qual uma única abertura é usada para ingerir alimentos e excretar resíduos, gerou amplo estudo e controvérsia sobre sua classificação. É um carnívoro que vive no fundo do mar, escavando e se alimentando de moluscos, provavelmente em suas formas larvais, em vez de adultos com conchas duras.
Sistemática
Etimologia
O nome do gênero Xenoturbella deriva do grego antigo xénos, que significa "estrangeiro" ou "estranho", e do latim turbela, que significa "agitação" ou "turbulência na água". O gênero Xenoturbella pertence ao subfilo Xenoturbellida, conhecido em sueco como paradoxmaskar,[2] ou "vermes paradoxais" (termo também usado por algumas mídias populares), pois, se classificado como deuterostômio, seria mais próximo dos humanos do que de outros invertebrados mais complexos, como lagostas.[3] Os deuterostômios formam um superfilo de animais cujo ânus se desenvolve antes da boca durante o desenvolvimento embrionário, incluindo humanos, outros cordados, equinodermos e hemicordados.
O epíteto específico bocki homenageia Sixten Bock, que coletou o organismo pela primeira vez em 1915.[4] O nome foi atribuído por Einar Westblad, que descreveu a espécie em 1949.
Taxonomia
Em 1999, a análise de espécimes de X. bocki armazenados no Museu Sueco de História Natural revelou que um pequeno subgrupo pertencia a outra espécie.[5] Esse grupo diferia dos espécimes identificados como X. bocki por apresentar fertilização interna, tamanho máximo de 12 mm e coloração rosa, em contraste com a coloração amarelo-branca de X. bocki. O novo táxon foi nomeado em homenagem a Westblad, que coletou os espécimes em habitats mais rasos e de sedimentos mais grossos na mesma região de X. bocki. No entanto, sequenciamento de DNA mitocondrial de espécimes das duas populações sugeriu que pertenciam à mesma espécie, indicando que X. westbladi é um sinônimo júnior de X. bocki.[6]
Filogenia
Nível de espécie
Comparações de sequências de DNA mitocondrial e proteínas mostraram que Xenoturbella bocki, frequentemente encontrada ao largo da costa da Suécia, é o grupo irmão de X. hollandorum, uma espécie descoberta em 2016 no leste do Oceano Pacífico.[7] Essas duas espécies compartilham afinidades evolutivas com X. japonica em um clado de táxons de "águas rasas".[8]
| Cladograma das espécies do género Xenoturbella. | |||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||
| O cladograma foi reconstruído a partir do DNA mitocondrial e das sequências proteicas.[7][8] |
Acima do nível de gênero
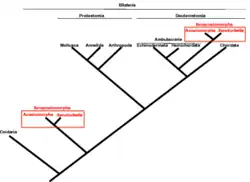
Quando descoberta, X. bocki foi colocada em um novo gênero, Xenoturbella. Acima do nível de gênero, sua classificação é controversa. Westblad a posicionou no filo Platyhelminthes, na classe Turbellaria (platelmintos de vida livre).[9] Em 1999, com base em análises genéticas, Israelsson a classificou como Protostomia, agrupada com bivalves.[10][11] Protostomia é um grande clado que inclui anelídeos, moluscos e artrópodes. No desenvolvimento embrionário, a boca se desenvolve antes do ânus na maioria dos protostômios, embora alguns apresentem caminhos de desenvolvimento alternativos. Se incluída neste clado, Xenoturbella também estaria entre essas exceções.[12]
No entanto, hoje essa classificação é considerada incorreta devido à contaminação por DNA de moluscos em sua dieta. Em 2006, a cientista sueca Sarah J. Bourlat e coautores a colocaram em seu próprio filo, Xenoturbellida. Estudos mais recentes, com base em evidências genéticas e de desenvolvimento (como genes Hox), sugerem que ela deve ser agrupada com Acoela e Nemertodermatida em Acoelomorpha. Esses três táxons são por vezes incluídos nos deuterostômios (um grande clado que abrange humanos, outros cordados, estrelas-do-mar e outros),[12] enquanto outros os classificam como um ramo basal que se assemelha a um ancestral comum de deuterostômios e protostômios.[11] Uma análise de 2016 de múltiplos conjuntos de dados genéticos apoia a última hipótese, sugerindo que, como Xenoturbella bocki, o ancestral comum de protostômios e deuterostômios provavelmente tinha uma abertura única, locomoção ciliada e corpo vermiforme.[13] Contudo, se a hipótese dos deuterostômios for correta, Xenoturbella deve ter perdido várias características ancestrais, como o ânus.[14]
Descrição
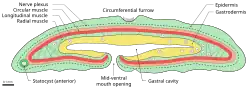
Este animal geralmente cresce até 1 cm de comprimento,[11] embora indivíduos de até 4 cm tenham sido registrados.[11] Seu sistema nervoso consiste em uma rede nervosa sem cérebro ou gânglios nervosos definidos. A rede nervosa está localizada no lado basal (afastado da superfície do animal) da pele.[15] Este animal não possui celoma. Também não tem ânus, excretando resíduos pela mesma abertura por onde ingere alimentos.[9] Assim, o órgão digestivo tem forma de saco. A abertura está localizada na região ventral, próxima à parte frontal. O animal é simultaneamente hermafrodita.[4]
Um sulco percorre a circunferência do corpo na região central do animal, além de sulcos laterais. Em seus lados, há numerosos cílios pequenos que auxiliam na locomoção. Pequenas células contêm vesículas que podem funcionar como glândulas. Um órgão de função desconhecida, preliminarmente chamado de estatocisto, foi observado na extremidade frontal do animal. Duas hipóteses principais sugerem que ele auxilia no equilíbrio, como os estatocistos em outros invertebrados, ou que possui funções endócrinas. Experimentos em que o animal foi observado se dividindo em dois após um ferimento mostram que o estatocisto é essencial para o comportamento normal e a sobrevivência a longo prazo.[9]
Ecologia
O animal se move na água por meio de contrações musculares rítmicas, auxiliadas pelos cílios laterais e por um tufo de cílios mais longos nas costas. O organismo também pode usar sua musculatura para se enrolar em uma bola, mantendo essa forma por vários meses.[9] Adultos mantêm uma relação simbiótica com Chlamydiae e Gammaproteobacteria, dois endossimbiontes bacterianos encontrados em sua gastroderme.[16] Dados genéticos confirmam que sua dieta inclui moluscos bivalves.[11] No entanto, nunca foi observado se alimentando, sendo desconhecido se consome carcaças de bivalves, ovos, esperma, muco, fezes ou bivalves larvais ou adultos vivos.[9] Não possui meios visíveis para penetrar as conchas de bivalves adultos. Espécimes em cativeiro sobreviveram por meses sem comida e não mostraram interesse nos itens alimentares propostos. Isso levou alguns a sugerirem que ele se alimenta absorvendo matéria orgânica dissolvida pela pele. Pelo menos um espécime, que supostamente mostra uma larva de bivalve consumida, está preservado no Museu Sueco de História Natural. Esta espécie escava, e foi observada criando túneis de até 15 cm de profundidade no substrato em aquários de laboratório.[9]
Distribuição
Esta espécie foi encontrada em habitats oceânicos ao largo da costa da Europa, mais frequentemente na costa da Suécia. É frequentemente coletada em lama do fundo do mar, a profundidades de 50 a 200 m.[9][17]
Reprodução
X. bocki só foi observada se reproduzindo sexualmente.[9] Na natureza, a espécie desova no inverno. Seus ovos, pequenos e revestidos de muco, afundam na coluna d'água.[11] Os ovos têm cor laranja-pálida e são opacos. Após a eclosão, os jovens são amarelados, quase esféricos, e movem-se para a superfície da água. As larvas não possuem blastóporo e não se alimentam até estarem completamente desenvolvidas, podendo obter nutrição do vitelo, o que as tornaria lecitotróficas. Em cinco dias, contrações musculares são observadas em laboratório, o que pode auxiliar na locomoção. X. bocki apresenta desenvolvimento direto. Até 2013, criar este animal em cativeiro era extremamente desafiador.[11]
Referências
- ↑ Westblad, E. (1949). «Xenoturbella bocki n. g., n. sp., a peculiar, primitive Turbellarian type». Arkiv för Zoologi. 1: 3–29
- ↑ «Xenoturbellida». Dyntaxa. Consultado em 31 de dezembro de 2018
- ↑ Jauregui, Andres (28 de março de 2013). «'Paradox Worm' Xenoturbella Bocki Lacks Brain & Sex Organs, But Could Be Mankind's 'Progenitor'». HuffPost. Consultado em 26 de abril de 2019
- ↑ a b Telford, Maximilian J. (2008). «Xenoturbellida: The fourth deuterostome phylum and the diet of worms». Genesis. 46 (11): 580–586. ISSN 1526-968X. PMID 18821586. doi:10.1002/dvg.20414

- ↑ Israelsson, Olle (22 de abril de 1999). «New light on the enigmatic Xenoturbella (phylum uncertain): ontogeny and phylogeny». Proceedings of the Royal Society of London. Series B, Biological Sciences. 266 (1421): 835–841. PMC 1689910. doi:10.1098/rspb.1999.0713
- ↑ Rouse, Greg W.; Wilson, Nerida G.; Carvajal, Jose I.; Vrijenhoek, Robert C. (2016). «New deep-sea species of Xenoturbella and the position of Xenacoelomorpha». Nature. 530 (7588): 94–97. Bibcode:2016Natur.530...94R. PMID 26842060. doi:10.1038/nature16545
- ↑ a b Rouse, Greg W.; Wilson, Nerida G.; Carvajal, Jose I.; Vrijenhoek, Robert C. (4 de fevereiro de 2016). «New deep-sea species of Xenoturbella and the position of Xenacoelomorpha». Nature (em inglês). 530 (7588): 94–97. Bibcode:2016Natur.530...94R. ISSN 0028-0836. PMID 26842060. doi:10.1038/nature16545
- ↑ a b Nakano, Hiroaki; Miyazawa, Hideyuki; Maeno, Akiteru; Shiroishi, Toshihiko; Kakui, Keiichi; Koyanagi, Ryo; Kanda, Miyuki; Satoh, Noriyuki; Omori, Akihito; Kohtsuka, Hisanori (18 de dezembro de 2017). «A new species of Xenoturbella from the western Pacific Ocean and the evolution of Xenoturbella». BMC Evolutionary Biology (em inglês). 17 (1). 245 páginas. ISSN 1471-2148. PMC 5733810. PMID 29249199. doi:10.1186/s12862-017-1080-2
- ↑ a b c d e f g h Nakano, Hiroaki (2015). «What is Xenoturbella?». Zoological Letters. 1 (22). 22 páginas. PMC 4657256. PMID 26605067. doi:10.1186/s40851-015-0018-z
- ↑ Perseke, Marleen; Hankeln, Thomas; Weich, Bettina; Fritzsch, Guido; Stadler, Peter F.; Israelsson, Olle; Bernhard, Detlef; Schlegel, Martin (abril de 2007). «The Mitochondrial DNA of Xenoturbella bocki: Genomic Architecture and Phylogenetic Analysis». Theory in Biosciences. 126 (1): 35–42. PMID 18087755. doi:10.1007/s12064-007-0007-7
- ↑ a b c d e f g Nakano, H.; Lundin, K.; Bourlat, S. J.; Telford, M. J.; Funch, P.; Nyengaard, J. R.; Obst, M.; Thorndyke, M. C. (2013). «Xenoturbella bocki exhibits direct development with similarities to Acoelomorpha». Nature Communications. 4: 1537. Bibcode:2013NatCo...4.1537N. PMC 3586728. PMID 23443565. doi:10.1038/ncomms2556
- ↑ a b de Mendoza, A.; Ruiz-Trillo, I. (2011). «The Mysterious Evolutionary Origin for the GNE Gene and the Root of Bilateria». Molecular Biology and Evolution. 28 (11): 2987–2991. PMC 4342547. PMID 21616910. doi:10.1093/molbev/msr142
- ↑ Cannon, J. T.; Vellutini, B. C.; Smith, J.; Ronquist, F.; Jondelius, U.; Hejnol, A. (2016). «Xenacoelomorpha is the sister group to Nephrozoa». Nature. 530 (7588): 89–93. Bibcode:2016Natur.530...89C. PMID 26842059. doi:10.1038/nature16520
- ↑ Philippe, H.; Brinkmann, H.; Copley, R. R.; Moroz, L. L.; Nakano, H.; Poustka, A. J.; Wallberg, A.; Peterson, K. J.; Telford, M. J. (2011). «Acoelomorph flatworms are deuterostomes related to Xenoturbella». Nature. 470 (7333): 255–258. Bibcode:2011Natur.470..255P. PMC 4025995. PMID 21307940. doi:10.1038/nature09676
- ↑ Schmidt-Rhaesa, Andreas; Steffen, Harzsch; Günter, Purschke; Thomas, Stach (2016). Structure and Evolution of Invertebrate Nervous Systems 1st ed. Oxford: Oxford University Press. ISBN 9780191066214
- ↑ Kjeldsen, K. U.; Obst, M.; Nakano, H.; Funch, P.; Schramm, A. (2010). «Two Types of Endosymbiotic Bacteria in the Enigmatic Marine Worm Xenoturbella bocki». Applied and Environmental Microbiology. 76 (8): 2657–2662. Bibcode:2010ApEnM..76.2657K. PMC 2849209. PMID 20139320. doi:10.1128/aem.01092-09
- ↑ Nakano, Hiroaki; Miyazawa, Hideyuki; Maeno, Akiteru; Shiroishi, Toshihiko; Kakui, Keiichi; Koyanagi, Ryo; Kanda, Miyuki; Satoh, Noriyuki; Omori, Akihito; Kohtsuka, Hisanori (2017). «A new species of Xenoturbella from the western Pacific Ocean and the evolution of Xenoturbella». BMC Evolutionary Biology. 17 (1): 245. PMC 5733810. PMID 29249199. doi:10.1186/s12862-017-1080-2
Ligações externas
 Media relacionados com Xenoturbella bocki no Wikimedia Commons
Media relacionados com Xenoturbella bocki no Wikimedia Commons