Voo de insetos

Insetos são o único grupo de invertebrados que desenvolveram asas e voo. Os insetos voaram pela primeira vez no Carbonífero, há cerca de 300 a 350 milhões de anos, tornando-se os primeiros animais a desenvolverem o voo. As asas podem ter evoluído a partir de apêndices nas laterais de membros existentes, que já possuíam nervos, articulações e músculos usados para outros fins. Inicialmente, esses apêndices podem ter sido usados para navegar na água ou para reduzir a velocidade de descida durante o planeio.
Dois grupos de insetos, as libélulas e os efemerópteros, possuem músculos de voo diretamente ligados às asas. Em outros insetos alados, os músculos de voo estão ligados ao tórax, que faz com que ele oscile para induzir o batimento das asas. Desses insetos, alguns (moscas e alguns besouros) alcançam frequências de batimento de asas muito altas através da evolução de um sistema nervoso "assíncrono", no qual o tórax oscila mais rápido que a taxa de impulsos nervosos.
Nem todos os insetos são capazes de voar. Alguns insetos ápteros perderam secundariamente suas asas através da evolução, enquanto outros insetos mais basais, como a traça Lepisma saccharina, nunca desenvolveram asas. Em alguns insetos eusociais, como formigas e cupins, apenas as castas reprodutivas desenvolvem asas durante a temporada de acasalamento antes de perdê-las após o acasalamento, enquanto os membros de outras castas permanecem sem asas durante toda a vida.
Alguns insetos muito pequenos utilizam não a aerodinâmica de estado estacionário, mas o mecanismo de batida e lançamento de Weis-Fogh, gerando grandes forças de sustentação ao custo de desgaste nas asas. Muitos insetos podem pairar, mantendo a altitude e controlando sua posição. Alguns insetos, como mariposas, têm as asas anteriores acopladas às asas posteriores para que funcionem em uníssono.
Mecanismos
Voo direto
Diferentemente de outros insetos, os músculos das asas das efemerópteros e Odonata (libélulas e donzelinhas) inserem-se diretamente nas bases das asas, que são articuladas de modo que um pequeno movimento descendente da base da asa eleva a própria asa para cima, semelhante a remar pelo ar. Libélulas e donzelinhas têm asas anteriores e posteriores semelhantes em forma e tamanho. Cada uma opera independentemente, proporcionando um grau de controle fino e mobilidade em termos da brusquidão com que podem mudar de direção e velocidade, algo não visto em outros insetos voadores. Odonatas são todos predadores aéreos, e eles sempre caçaram outros insetos voadores.[1]
-
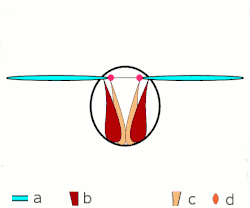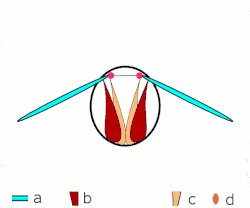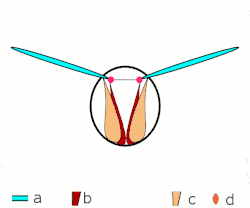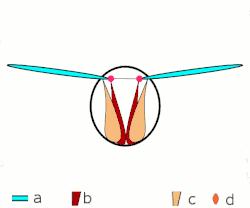 Voo direto: músculos ligados às asas. Apenas em insetos grandes.
Voo direto: músculos ligados às asas. Apenas em insetos grandes. -
 Os Odonata (libélulas e donzelinhas) têm musculatura de voo direto, assim como os efemerópteros.
Os Odonata (libélulas e donzelinhas) têm musculatura de voo direto, assim como os efemerópteros. -
Voo de libélula em câmera lenta.
Voo indireto
Diferentemente das duas ordens com músculos de voo direto, todos os outros insetos alados vivos voam usando um mecanismo diferente, envolvendo músculos de voo indireto. Esse mecanismo evoluiu uma vez e é a característica definidora (sinapomorfia) para a infraclasse Neoptera; ele corresponde, provavelmente não por coincidência, com o aparecimento de um mecanismo de dobramento de asas, que permite que insetos neópteros dobrem as asas sobre o abdômen quando em repouso (embora essa habilidade tenha sido perdida secundariamente em alguns grupos, como nas borboletas).[1]
O que todos os Neoptera compartilham, no entanto, é a forma como os músculos no tórax funcionam: esses músculos, em vez de se fixarem nas asas, fixam-se no tórax e deformam-no; como as asas são extensões do exoesqueleto torácico, as deformações do tórax causam o movimento das asas. Um conjunto de músculos longitudinais ao longo das costas comprime o tórax de frente para trás, fazendo com que a superfície dorsal do tórax (noto) se curve para cima, fazendo as asas descerem. Outro conjunto de músculos do tergo ao esterno puxa o noto para baixo novamente, fazendo as asas subirem.[1][2]
-
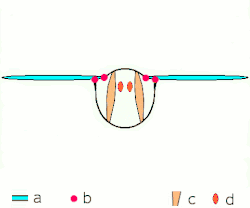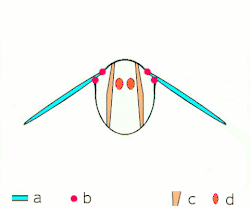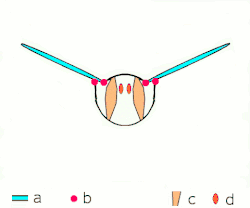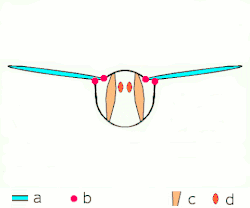 Voo indireto: músculos oscilam o tórax.
Voo indireto: músculos oscilam o tórax.
a: asas; b: articulações primárias/secundárias de voo; c: músculos de voo dorsoventrais; d: músculos longitudinais. -
 Os Neoptera, incluindo borboletas e a maioria dos outros insetos, têm musculatura de voo indireto.
Os Neoptera, incluindo borboletas e a maioria dos outros insetos, têm musculatura de voo indireto.
Insetos que batem suas asas menos de cem vezes por segundo usam músculo síncrono. Músculo síncrono é um tipo de músculo que contrai uma vez para cada impulso nervoso. Isso geralmente produz menos potência e é menos eficiente que o músculo assíncrono, o que explica a evolução independente de músculos de voo assíncronos em vários clados de insetos.[3]
Insetos que batem suas asas mais rapidamente, como as abelhas mamangavas, usam músculo assíncrono; este é um tipo de músculo que contrai mais de uma vez por impulso nervoso. Isso é alcançado estimulando o músculo a contrair novamente pela liberação de tensão no músculo, o que pode acontecer mais rapidamente do que através da estimulação nervosa simples.[4] Isso permite que a frequência de batimento de asas exceda a taxa na qual o sistema nervoso pode enviar impulsos. O músculo assíncrono é um dos últimos refinamentos que apareceram em alguns dos Neoptera superiores (Coleoptera, Diptera, e Hymenoptera). O efeito geral é que muitos Neoptera superiores podem bater suas asas muito mais rápido que insetos com músculos de voo direto.[1]
O músculo assíncrono está, por definição, sob controle relativamente grosseiro pelo sistema nervoso. Para equilibrar essa troca evolutiva, insetos que evoluíram com o voo indireto também desenvolveram um sistema neuromuscular separado para controle fino do movimento das asas.[5] Conhecidos como "músculos diretos", esses músculos fixam-se diretamente às escleras que compõem a articulação da asa e são contraídos com impulsos 1:1.[6] Trabalhos recentes começaram a abordar a dinâmica muscular complexa não linear na articulação da asa e seus efeitos no caminho da ponta da asa.[7]
Aerodinâmica
Existem dois modelos aerodinâmicos básicos de voo de insetos: criação de um vórtice de bordo de ataque e uso de batida e lançamento.[8][9]
Vórtice de bordo de ataque
A maioria dos insetos usa um método que cria um vórtice de bordo de ataque em espiral. Essas asas batem através de dois movimentos básicos de meia batida. O movimento descendente começa para cima e para trás e é mergulhado para baixo e para frente. Em seguida, a asa é rapidamente virada (supinação) de modo que o bordo de ataque aponta para trás. O movimento ascendente então empurra a asa para cima e para trás. Em seguida, a asa é virada novamente (pronação) e outro movimento descendente pode ocorrer. A faixa de frequência em insetos com músculos de voo síncronos é tipicamente de 5 a 200 hertz (Hz). Em aqueles com músculos de voo assíncronos, a frequência de batimento de asas pode exceder 1000 Hz. Quando o inseto está pairando, os dois movimentos levam o mesmo tempo. Um movimento descendente mais lento, no entanto, fornece impulso.[8][9]
A identificação de forças principais é crítica para entender o voo de insetos. As primeiras tentativas de entender asas batendo assumiram um estado quase estacionário. Isso significa que o fluxo de ar sobre a asa em qualquer momento dado foi assumido como sendo o mesmo que o fluxo seria sobre uma asa de estado estacionário não batendo no mesmo ângulo de ataque. Ao dividir a asa batendo em um grande número de posições imóveis e depois analisar cada posição, seria possível criar uma linha do tempo das forças instantâneas na asa a cada momento. O levantamento calculado foi considerado pequeno demais por um fator de três, então os pesquisadores perceberam que deve haver fenômenos instáveis fornecendo forças aerodinâmicas. Alguns pesquisadores previram picos de força na supinação. Com um modelo em escala dinâmica de uma mosca-das-frutas, essas forças previstas foram posteriormente confirmadas. Outros argumentaram que os picos de força durante a supinação e pronação são causados por um efeito rotacional desconhecido que é fundamentalmente diferente dos fenômenos translacionais. Há algum desacordo com esse argumento. Através de dinâmica dos fluidos computacional, alguns pesquisadores argumentam que não há efeito rotacional. Eles afirmam que as altas forças são causadas por uma interação com o rastro derramado pelo movimento anterior.[8][9]
Semelhante ao efeito rotacional mencionado acima, os fenômenos associados às asas batendo não são completamente compreendidos ou aceitos. Como cada modelo é uma aproximação, diferentes modelos deixam de fora efeitos que são presumidos como negligenciáveis. Por exemplo, o efeito Wagner, conforme proposto por Herbert A. Wagner em 1925,[10] diz que a circulação sobe lentamente para seu estado estacionário devido à viscosidade quando uma asa inclinada é acelerada a partir do repouso. Esse fenômeno explicaria um valor de sustentação menor do que o previsto. Normalmente, o caso foi para encontrar fontes para a sustentação adicional. Tem-se argumentado que esse efeito é negligenciável para o fluxo com um número de Reynolds típico do voo de insetos. O número de Reynolds é uma medida de turbulência; o escoamento laminar (suave) ocorre quando o número de Reynolds é baixo, e turbulento quando é alto.[11] O efeito Wagner foi ignorado, conscientemente, em pelo menos um modelo.[9]
Um dos fenômenos mais importantes que ocorre durante o voo de insetos é a sucção do bordo de ataque. Esse fenômeno foi proposto pela primeira vez por D. G. Ellis e J. L. Stollery em 1988 para descrever a sustentação de vórtices em asas delta de bordas agudas.[12] Em altos ângulos de ataque, o fluxo se separa sobre o bordo de ataque, mas se reanexa antes de alcançar a borda de fuga. Dentro dessa bolha de fluxo separado, há um vórtice. Como o ângulo de ataque é tão alto, muito momentum é transferido para baixo no fluxo. Esses dois recursos criam uma grande quantidade de força de sustentação, bem como algum arrasto adicional. Esse fenômeno é chamado atraso de estol, percebido pela primeira vez em hélices de aeronaves por H. Himmelskamp em 1945.[13] Esse efeito foi observado no voo de insetos batendo e foi comprovado ser capaz de fornecer sustentação suficiente para explicar a deficiência nos modelos de estado quase estacionário. Esse efeito é usado por remadores em um tipo de remada de puxada.[8][9]
Todos os efeitos em uma asa batendo podem ser reduzidos a três fontes aerodinâmicas principais: o vórtice de bordo de ataque, as forças aerodinâmicas de estado estacionário na asa e o contato da asa com seu rastro de movimentos anteriores. O tamanho dos insetos voadores varia de cerca de 20 microgramas a cerca de 3 gramas. À medida que a massa corporal do inseto aumenta, a área da asa aumenta e a frequência de batimento de asas diminui. Para insetos maiores, o número de Reynolds (Re) pode ser tão alto quanto 10000, onde o fluxo está começando a se tornar turbulento. Para insetos menores, pode ser tão baixo quanto 10. Isso significa que os efeitos viscosos são muito mais importantes para os insetos menores.[9][14][15]
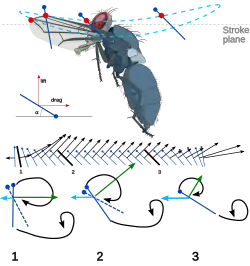
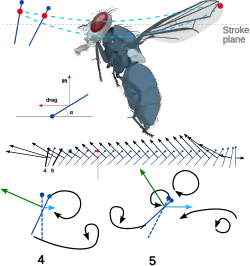
Outra característica interessante do voo dos insetos é a inclinação do corpo. À medida que a velocidade de voo aumenta, o corpo do inseto tende a se inclinar para baixo e se tornar mais horizontal. Isso reduz a área frontal e, portanto, a resistência do corpo. Como o arrasto também aumenta à medida que a velocidade de avanço aumenta, o inseto está tornando seu voo mais eficiente à medida que essa eficiência se torna mais necessária. Além disso, ao alterar o ângulo geométrico de ataque no curso descendente, o inseto consegue manter seu voo em uma eficiência ideal durante o maior número possível de manobras. O desenvolvimento do impulso geral é relativamente pequeno em comparação com as forças de elevação. As forças de elevação podem ser mais de três vezes o peso do inseto, enquanto o empuxo, mesmo nas velocidades mais altas, pode ser tão baixo quanto 20% do peso. Essa força é desenvolvida principalmente por meio do movimento ascendente menos potente do movimento de batimento ("flapping motion").[9][16]
Batida e lançamento


O mecanismo de batida e lançamento, ou mecanismo de Weis-Fogh, descoberto pelo zoólogo dinamarquês Torkel Weis-Fogh [en], é um método de geração de sustentação utilizado durante o voo de pequenos insetos.[17] Quando o tamanho dos insetos se torna menor que 1 mm, as forças viscosas tornam-se dominantes e a eficácia da geração de sustentação a partir de uma asa diminui drasticamente. Partindo da posição de batida, as duas asas se separam e giram em torno do bordo de fuga. As asas então se separam e "varrem" horizontalmente até o final do movimento descendente. Em seguida, as asas pronam e utilizam o bordo de ataque durante um movimento ascendente de remada. À medida que o movimento de batida começa, os bordos de ataque se encontram e giram juntas até que a lacuna desapareça. Inicialmente, pensava-se que as asas estavam se tocando, mas vários incidentes indicam uma lacuna entre as asas e sugerem que ela proporciona um benefício aerodinâmico.[18][19]
A geração de sustentação a partir do mecanismo de batida e lançamento ocorre durante vários processos ao longo do movimento. Primeiro, o mecanismo depende de uma interação asa-asa, já que o movimento de uma única asa não produz sustentação suficiente.[20][21][22] À medida que as asas giram em torno do bordo de fuga no movimento de lançamento, o ar corre para dentro da lacuna criada e gera um forte vórtice de bordo de ataque, e um segundo se desenvolve nas pontas das asas. Um terceiro vórtice, mais fraco, se desenvolve no bordo de fuga.[19] A força dos vórtices em desenvolvimento depende, em parte, da lacuna inicial de separação entre asas no início do movimento de lançamento. Um movimento de elevação durante o lançamento,[23] asas flexíveis,[21] e um mecanismo de atraso de estol foram encontrados para reforçar a estabilidade e fixação do vórtice.[24] Finalmente, para compensar a menor produção de sustentação durante o voo com baixos números de Reynolds (com escoamento laminar), insetos minúsculos frequentemente têm uma maior frequência de batimento para gerar velocidades de ponta de asa comparáveis às de insetos maiores.[24]
O maior arrasto esperado ocorre durante o movimento de lançamento dorsal, pois as asas precisam se separar e girar.[22] A atenuação das grandes forças de arrasto ocorre por meio de vários mecanismos. Asas flexíveis foram encontradas para diminuir o arrasto no movimento de lançamento em até 50% e reduzir ainda mais o arrasto total ao longo de todo o curso da asa quando comparado com asas rígidas.[21] Cerdas nas bordas das asas, como visto em Encarsia formosa, causam uma porosidade no fluxo que aumenta e reduz as forças de arrasto, ao custo de uma menor geração de sustentação.[25] Além disso, a separação entre as asas antes do lançamento desempenha um papel importante no efeito geral do arrasto. À medida que a distância entre as asas aumenta, o arrasto geral diminui.[18]
O mecanismo de batida e lançamento também é empregado pelo molusco marinho Limacina helicina, uma borboleta-do-mar.[26][27] Alguns insetos, como a mosca minadora de folhas Liriomyza sativae, exploram um mecanismo parcial de batida e lançamento, usando o mecanismo apenas na parte externa da asa para aumentar a sustentação em cerca de 7% quando pairando.[24]
- Mecanismo de batida e lançamento após Sane 2003
-
 Batida 1: asas fecham sobre as costas.
Batida 1: asas fecham sobre as costas. -
 Batida 2: bordos de ataque se tocam, asa gira em torno do bordo de ataque, vórtices se formam.
Batida 2: bordos de ataque se tocam, asa gira em torno do bordo de ataque, vórtices se formam. -
 Batida 3: bordos de fuga se fecham, vórtices são liberados, asas se fecham dando impulso.
Batida 3: bordos de fuga se fecham, vórtices são liberados, asas se fecham dando impulso.
- Setas pretas (curvadas): fluxo; Setas azuis: velocidade induzida; Setas laranjas: força líquida na asa.
-
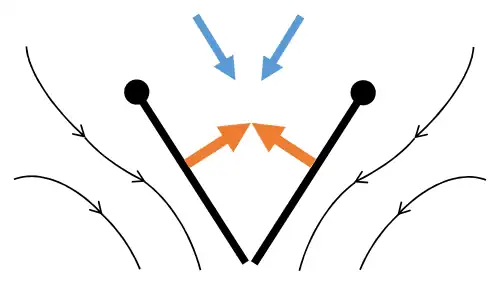 Lançamento 1: asas giram em torno do bordo de fuga para se separarem.
Lançamento 1: asas giram em torno do bordo de fuga para se separarem. -
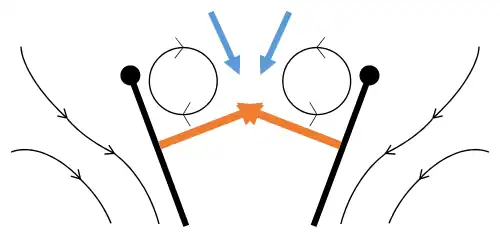 Lançamento 2: bordo de ataque se afasta, o ar corre para dentro, aumentando a sustentação.
Lançamento 2: bordo de ataque se afasta, o ar corre para dentro, aumentando a sustentação. -
 Lançamento 3: novo vórtice se forma no bordo de ataque, vórtices do bordo de fuga se cancelam, talvez ajudando o fluxo a crescer mais rápido.
Lançamento 3: novo vórtice se forma no bordo de ataque, vórtices do bordo de fuga se cancelam, talvez ajudando o fluxo a crescer mais rápido.
Equações governantes
Uma asa movendo-se em fluidos experimenta uma força fluida, que segue as convenções encontradas na aerodinâmica. O componente de força normal à direção do fluxo relativo à asa é chamado de sustentação (L - "lift"), e o componente de força na direção oposta ao fluxo é o arrasto (D - "drag"). Nos números de Reynolds considerados aqui, uma unidade de força apropriada é 1/2(ρU2S), onde ρ é a densidade do fluido, S a área da asa, e U a velocidade da asa. As forças adimensionais são chamadas de coeficientes de sustentação (CL) e arrasto (CD) , isso é:[8]

CL e CD são constantes apenas se o fluxo for estacionário. Uma classe especial de objetos como aerofólios pode atingir um estado estacionário quando corta o fluido em um pequeno ângulo de ataque. Nesse caso, o escoamento invíscido ao redor de um aerofólio pode ser aproximado por um fluxo potencial satisfazendo a condição de fronteira sem penetração. O teorema de Kutta Joukowski de um aerofólio 2D assume ainda que o fluxo deixa a borda de fuga acentuada suavemente, e isso determina a circulação total ao redor de um aerofólio. O levantamento correspondente é dado pelo teorema de Bernoulli (teorema de Blasius):[8]

Os fluxos ao redor de pássaros e insetos podem ser considerados incompressíveis: O número de Mach, ou velocidade relativa à velocidade do som no ar, é tipicamente 1/300 e a frequência da asa é cerca de 10–103 Hz. Usando a equação governante como uma das equações de Navier–Stokes sujeita à condição de fronteira sem deslizamento, a equação é:[8]

Onde u(x, t) é o campo de fluxo, p a pressão, ρ a densidade do fluido, ν a viscosidade cinemática, ubd a velocidade na fronteira, e us a velocidade do sólido. Ao escolher uma escala de comprimento, L, e escala de velocidade, U, a equação pode ser expressa em forma não dimensional contendo o número de Reynolds, Re=uL/ν . Existem duas diferenças óbvias entre uma asa de inseto e um aerofólio: uma asa de inseto é muito menor e ela bate. Usando uma libélula como exemplo, seu acorde (c) é cerca de 1 cm, seu comprimento de asa (l) cerca de 4 cm, e sua frequência de asa (f) cerca de 40 Hz. A velocidade da ponta (u) é cerca de 1 m/s, e o número de Reynolds correspondente é cerca de 103. No extremo menor, uma vespa da superfamília Chalcidoidea típica tem um comprimento de asa de cerca de 0.5-0.7 mm e bate sua asa a cerca de 400 Hz. A faixa do número de Reynolds no voo de insetos é de cerca de 10 a 104, que está entre os dois limites convenientes para as teorias: escoamentos estacionários não viscosos ao redor de um aerofólio e escoamento de Stokes experimentado por uma bactéria nadadora. Por essa razão, esse intervalo intermediário não é bem compreendido. Por outro lado, é talvez o regime mais onipresente entre as coisas que vemos. Folhas e sementes que caem, peixes e pássaros encontram escoamentos instáveis semelhantes aos vistos ao redor de um inseto.[8] O número de Reynolds cordal pode ser descrito por:

e


Onde é o comprimento médio do acorde, é a velocidade da ponta da asa, é a amplitude do batimento, é a frequência de batimento, é o raio de giração, é a área da asa, e é o comprimento da asa, incluindo a ponta da asa.







Além do número de Reynolds, existem pelo menos dois outros parâmetros adimensionais relevantes. Uma asa tem três escalas de velocidade: a velocidade de batimento em relação ao corpo (u), a velocidade de avanço do corpo (U0), e a velocidade de inclinação (Ωc). As proporções delas formam duas variáveis adimensionais, U0/u e Ωc/u, a primeira é frequentemente referida como a razão de avanço, e está também relacionada à frequência reduzida, fc/U0.[8]
Se uma asa de inseto for rígida, por exemplo, uma asa de Drosophila é aproximadamente assim, seu movimento em relação a um corpo fixo pode ser descrito por três variáveis: a posição da ponta em coordenadas esféricas, (Θ(t),Φ(t)), e o ângulo de inclinação ψ(t), em torno do eixo que conecta a raiz e a ponta. Para estimar as forças aerodinâmicas com base na análise do elemento da lâmina, também é necessário determinar o ângulo de ataque (α). O ângulo de ataque típico a 70% da envergadura varia de 25° a 45° em insetos pairando (15° em beija-flores). Apesar da riqueza de dados disponíveis para muitos insetos, relativamente poucos experimentos relatam a variação temporal de α durante um batimento. Entre eles estão experimentos em túneis de vento de uma vespa amarrada e uma mosca amarrada, e voo livre pairando de uma mosca-das-frutas.[8]
Porque são relativamente fáceis de medir, as trajetórias das pontas das asas foram relatadas com mais frequência. Por exemplo, selecionando apenas sequências de voo que produziram sustentação suficiente para suportar um peso, mostrará que a ponta da asa segue uma forma elíptica. Formas não cruzadas também foram relatadas para outros insetos. Independentemente de suas formas exatas, o movimento de mergulho indica que os insetos podem usar arrasto aerodinâmico além da sustentação para suportar seu peso.[8]
Pairar
| Parâmetros de voo[28] | Velocidade (m/s) | Batidas/s |
|---|---|---|
| Libélulas Aeshnidae [en]. | 7.0 | 38 |
| Vespas | 5.7 | 100 |
| Mariposa-esfinge-colibri | 5.0 | 85 |
| Mutucas | 3.9 | 96 |
| Moscas-das-flores | 3.5 | 120 |
| Mamangavas | 2.9 | 130 |
| Abelhas Apis | 2.5 | 250 |
| Moscas-domésticas | 2.0 | 190 |
| Donzelinhas | 1.5 | 16 |
| Mecoptera | 0.49 | 28 |
| Borboleta branca da couve[29] | 2.5 | 12 |
| Tripes[24] | 0.3 | 254 |
.jpg)
Muitos insetos podem pairar, ou permanecer em um ponto no ar, fazendo isso ao baterem suas asas rapidamente. Fazê-lo requer estabilização lateral, bem como a produção de sustentação. A força de sustentação é produzida principalmente pelo movimento descendente. Como as asas empurram o ar ao redor, a força de reação do ar nas asas empurra o inseto para cima. As asas da maioria dos insetos evoluíram de modo que, durante o movimento ascendente, a força na asa é pequena. Como o batimento descendente e o retorno forçam o inseto para cima e para baixo, respectivamente, o inseto oscila e acaba permanecendo na mesma posição.[14]
A distância que o inseto cai entre os batimentos de asas depende de quão rapidamente suas asas estão batendo: quanto mais lento ele bate, maior o intervalo em que ele cai, e maior a distância que ele cai entre cada batimento de asas. Pode-se calcular a frequência de batimento de asas necessária para o inseto manter uma dada estabilidade em sua amplitude. Para simplificar os cálculos, deve-se assumir que a força de sustentação é um valor constante finito enquanto as asas estão se movendo para baixo e que é zero enquanto as asas estão se movendo para cima. Durante o intervalo de tempo Δt do batimento ascendente, o inseto cai uma distância h sob a influência da gravidade.[14]

O movimento ascendente então restaura o inseto à sua posição original. Tipicamente, pode ser necessário que a posição vertical do inseto não mude mais que 0,1 mm (ou seja, h = 0,1 mm). O tempo máximo permitido para a queda livre é então[14]

Como os movimentos para cima e para baixo das asas têm aproximadamente a mesma duração, o período T para um ciclo completo de batimento de asas é o dobro de Δr, ou seja,[14]

A frequência dos batimentos, f, significando o número de batimentos de asas por segundo, é representada pela equação:[14]

No exemplo usado, a frequência é de 110 batimentos/s, que é a frequência típica encontrada em insetos. Borboletas têm uma frequência muito menor com cerca de 10 batimentos/s, o que significa que elas não podem pairar. Outros insetos podem ser capazes de produzir uma frequência de 1000 batimentos/s. Para restaurar o inseto à sua posição vertical original, a força média ascendente durante o movimento descendente, Fav, deve ser igual a duas vezes o peso do inseto. Observe que, como a força ascendente no corpo do inseto é aplicada apenas por metade do tempo, a força média ascendente no inseto é simplesmente seu peso.[14]
Entrada de energia
Pode-se agora calcular a energia necessária para o inseto se manter pairando, considerando novamente um inseto com massa de 0,1 g, força média, Fav, aplicada pelas duas asas durante o movimento descendente é duas vezes o peso. Como a pressão aplicada pelas asas é uniformemente distribuída sobre a área total da asa, isso significa que pode-se assumir que a força gerada por cada asa atua através de um único ponto na seção média das asas. Durante o movimento descendente, o centro das asas percorre uma distância vertical d.[14] O trabalho ("work") total realizado pelo inseto durante cada golpe para baixo é o produto da força e da distância, ou seja:

Se as asas varrem através do batimento em um ângulo de 70°, então, no caso apresentado para o inseto com asas de 1 cm de comprimento, d é 0,57 cm. Portanto, o trabalho realizado durante cada batimento pelas duas asas é:[14]

A energia E é usada para elevar o inseto contra a gravidade. A energia necessária para elevar a massa do inseto em 0,1 mm durante cada batida descendente é:[14]

Esta é uma fração insignificante da energia total gasta, o que indica claramente que a maior parte da energia é gasta em outros processos. Uma análise mais detalhada do problema mostra que o trabalho realizado pelas asas é convertido principalmente em energia cinética do ar que é acelerado pelo movimento descendente das asas. A potência é a quantidade de trabalho realizada em 1 s; no inseto usado como exemplo, faz 110 movimentos descendentes por segundo. Portanto, sua saída de potência P é, batimentos por segundo, e isso significa que sua saída de potência P é:[14]

Saída de potência
Na cálculo da potência usada no pairar dos insetos, os exemplos utilizados negligenciaram a energia cinética das asas em movimento. As asas dos insetos, leves como são, têm uma massa finita; portanto, como se movem, possuem energia cinética. Como as asas estão em movimento rotativo, a energia cinética máxima durante cada curso da asa é:[14]

Aqui I é o momento de inércia da asa e ωmax é a velocidade angular máxima durante o curso da asa. Para obter o momento de inércia para a asa, assumiremos que a asa pode ser aproximada por uma haste fina articulada em uma extremidade. O momento de inércia para a asa é então:[14]

Onde l é o comprimento da asa (1 cm) e m é a massa de duas asas, que pode ser tipicamente 10−3 g. A velocidade angular máxima, ωmax, pode ser calculada a partir da velocidade linear máxima, νmax, no centro da asa:[14]

Durante cada curso, o centro das asas se move com uma velocidade linear média νav dada pela distância d percorrida pelo centro da asa dividida pela duração Δt do curso da asa. A partir do nosso exemplo anterior, d = 0,57 cm e Δt = 4,5×10−3 s. Portanto:[14]

A velocidade das asas é zero tanto no início quanto no final do curso da asa, o que significa que a velocidade linear máxima é maior que a velocidade média. Se assumirmos que a velocidade oscila (de forma sinusoide) ao longo do caminho da asa, a velocidade linear máxima é duas vezes maior que a velocidade média. Portanto, a velocidade angular máxima é:[14]

E a energia cinética é:[14]

Como há dois cursos de asas (o movimento ascendente e descendente) em cada ciclo do movimento da asa, a energia cinética é 2×43 = 86 erg. Isso é aproximadamente a mesma quantidade de energia consumida no próprio pairar.[14]
Elasticidade

Os insetos ganham energia cinética, fornecida pelos músculos, quando as asas aceleram. Quando as asas começam a desacelerar em direção ao final do curso, essa energia deve se dissipar. Durante o movimento descendente, a energia cinética é dissipada pelos próprios músculos e é convertida em calor (esse calor é às vezes usado para manter a temperatura corporal central). Alguns insetos conseguem utilizar a energia cinética no movimento ascendente das asas para ajudar em seu voo. As articulações das asas desses insetos contêm uma almofada de proteína elástica, semelhante a borracha, chamada resilina [en]. Durante o movimento ascendente da asa, a resilina é esticada. A energia cinética da asa é convertida em energia potencial na resilina esticada, que armazena a energia como uma mola. Quando a asa desce, essa energia é liberada e auxilia no movimento descendente.[14]
Usando algumas suposições simplificadoras, podemos calcular a quantidade de energia armazenada na resilina esticada. Embora a resilina seja dobrada em uma forma complexa, o exemplo dado mostra o cálculo como uma haste reta de área A e comprimento. Além disso, assumiremos que ao longo do estiramento, a resilina obedece à lei de Hooke. Isso não é estritamente verdadeiro, pois a resilina é esticada em uma quantidade considerável e, portanto, tanto a área quanto o módulo de Young mudam no processo de esticamento. A energia potencial U armazenada na resilina esticada é:[14]

Aqui E é o módulo de Young para resilina, que foi medido como 1,8×107 dyn/cm2. Tipicamente, em um inseto do tamanho de uma abelha, o volume da resilina pode ser equivalente a um cilindro de 2×10−2 cm de comprimento e 4×10−4 cm2 de área. No exemplo dado, o comprimento da haste de resilina é aumentado em 50% quando esticado. Portanto, neste caso, a energia potencial armazenada na resilina de cada asa é:[14]

A energia armazenada nas duas asas para um inseto do tamanho de uma abelha é de 36 erg, o que é comparável à energia cinética no movimento ascendente das asas. Experimentos mostram que até 80% da energia cinética da asa pode ser armazenada na resilina.[14]
Acoplamento de asas
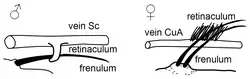
Algumas ordens de insetos, como os Lepidoptera, desenvolveram mecanismos morfológicos de acoplamento de asas [en] no imago que tornam esses taxa funcionalmente como de duas asas.[30] Todas as formas, exceto as mais basais, exibem esse acoplamento de asas.[31]
Há três tipos de mecanismos – jugal, frênulo-retinacular e amplexiforme:[32]
- Os grupos mais primitivos têm uma área lobular aumentada perto da margem posterior basal, ou seja, na base da asa anterior, um jugum, que se dobra sob a asa posterior em voo.[31][33]
- Outros grupos têm um frênulo na asa posterior que se engancha sob um retináculo na asa anterior.[33]
- Em quase todas as borboletas e nas mariposas Bombycoidea [en] (exceto as Sphingidae), não há um arranjo de frênulo e retináculo para acoplar as asas. Em vez disso, uma área humeral aumentada da asa posterior é amplamente sobreposta pela asa anterior. Apesar da ausência de uma conexão mecânica específica, as asas se sobrepõem e operam em fase. O movimento de força da asa anterior empurra a asa posterior em uníssono. Esse tipo de acoplamento é uma variação do tipo frenado, mas onde o frênulo e o retináculo estão completamente perdidos.[30][34]
Bioquímica
A bioquímica do voo de insetos tem sido um foco de estudo considerável. Enquanto muitos insetos usam carboidratos e lipídios como fonte de energia para o voo, muitos besouros e moscas usam o aminoácido prolina como sua fonte de energia.[35] Algumas espécies também usam uma combinação de fontes e mariposas como a Manduca sexta usam carboidratos para aquecimento antes do voo.[36]
Feedback sensorial
Os insetos usam feedback sensorial para manter e controlar o voo. Os insetos controlam a postura de voo, amplitude de batimento de asas e frequência de batimento usando informações de suas antenas,[37] halteres,[38] e asas.[39]
Evolução e adaptação
Em algum momento do período Carbonífero, há cerca de 350 a 300 milhões de anos, quando havia apenas duas grandes massas de terra, os insetos começaram a voar.[40] Entre os fósseis de insetos alados mais antigos está Delitzschala, um Palaeodictyoptera do Baixo Carbonífero;[41] e Rhyniognatha [en] que é mais antigo, do Devoniano Inferior, mas não está claro se ele tinha asas, ou mesmo se era um inseto.[42][43]
Como e por que as asas dos insetos se desenvolveram não é bem compreendido, em grande parte devido à escassez de fósseis apropriados do período de seu desenvolvimento no Baixo Carbonífero. Historicamente, havia três teorias principais sobre a origem das asas dos insetos. A primeira era que elas são modificações de brânquias abdominais móveis, como encontradas nas ninfas aquáticas de efémeras. Análises filogenômicas sugerem que os Polyneoptera, o grupo de insetos alados que inclui gafanhotos, evoluíram de um ancestral terrestre, tornando a evolução das asas a partir de brânquias improvável.[44] Estudos adicionais sobre o comportamento de salto de larvas de efémeras determinaram que as brânquias traqueais não desempenham um papel na condução da descida de insetos, fornecendo mais evidências contra essa hipótese evolutiva.[45] Isso deixa duas grandes teorias históricas: que as asas se desenvolveram a partir de lobos paranotais, extensões do tergo torácico; ou que elas surgiram de modificações de segmentos de pernas, que já continham músculos.[46]
Hipótese epicoxal (brânquias abdominais)
.JPG)
Numerosos entomologistas,[47] incluindo Hermann Landois [en] em 1871, Lubbock em 1873, Vitus Graber [en] em 1877 e Herbert Osborn [en] em 1905 sugeriram que uma possível origem para as asas de insetos poderia ser brânquias abdominais móveis encontradas em muitos insetos aquáticos, como nas ninfas de efémeras. De acordo com essa teoria, essas brânquias traqueais, que começaram como saídas do sistema respiratório e com o tempo foram modificadas para propósitos locomotores, eventualmente se desenvolveram em asas. As brânquias traqueais estão equipadas com pequenas asas que vibram perpetuamente e têm seus próprios músculos retos minúsculos.[48]
Hipótese paranotal (tergal)
A hipótese do lobo paranotal ou tergal (parede dorsal do corpo), proposta por Fritz Müller em 1875[49] e reformulada por G. Crampton em 1916,[47] Jarmila Kukalova-Peck em 1978[50] e Alexander P. Rasnitsyn em 1981 entre outros,[51] sugere que as asas dos insetos se desenvolveram a partir de lobos paranotais, uma preadaptação encontrada em fósseis de insetos que teriam auxiliado na estabilização enquanto pulavam ou caíam. A favor dessa hipótese está a tendência de muitos insetos, quando assustados enquanto escalam galhos, escaparem caindo no chão. Esses lobos teriam servido como paraquedas e possibilitariam que o inseto aterrissasse mais suavemente. A teoria sugere que esses lobos gradualmente aumentaram de tamanho e em um estágio posterior desenvolveram uma articulação com o tórax. Mesmo mais tarde, apareceriam os músculos para mover essas asas rudimentares. Este modelo implica um aumento progressivo na eficácia das asas, começando com paraquedismo, depois planagem e finalmente voo ativo. Ainda assim, a falta de evidências fósseis substanciais do desenvolvimento das articulações e músculos das asas representa uma grande dificuldade para a teoria, assim como o desenvolvimento aparentemente espontâneo de articulação e venação, e ela foi amplamente rejeitada por especialistas na área.[48]
Hipótese endite-exite (pleural)

Em 1990, J. W. H. Trueman propôs que a asa foi adaptada dos "enditos" e "exitos", apêndices nos respectivas estruturas internas e externas do membro do artrópode primitivo, também chamada de hipótese pleural. Isso se baseou em um estudo realizado por Goldschmidt em 1945 em Drosophila melanogaster, no qual uma variação chamada "pod" (para podômeros, segmentos do membro) apresentava uma mutação que transformava as asas normais. O resultado foi interpretado como um arranjo de pernas triplamente articuladas com alguns apêndices adicionais, mas sem o tarso, onde normalmente estaria a superfície costal da asa. Essa mutação foi reinterpretada como uma forte evidência de uma fusão do exito e do endito dorsal, em vez de uma perna, com os apêndices se encaixando muito melhor nessa hipótese. A inervação, a articulação e a musculatura necessárias para a evolução das asas já estão presentes nos segmentos dos membros.[46]
Outras hipóteses
Outras hipóteses incluem a sugestão de Vincent Wigglesworth, de 1973, de que as asas se desenvolveram a partir de protuberâncias torácicas usadas como radiadores.[48] Adrian Thomas e Åke Norberg sugeriram em 2003 que as asas podem ter se desenvolvido inicialmente para navegar na superfície da água, como observado em alguns Plecoptera.[52][53] Stephen P. Yanoviak e seus colegas propuseram em 2009 que a asa deriva de uma descida aérea planar direcionada - um fenômeno pré-voo encontrado em alguns apterigotos, um táxon irmão sem asas dos insetos alados.[54]
Origem dupla
Biólogos como Averof,[55] Niwa,[56] Elias-Neto[57] e seus colegas começaram a explorar a origem da asa dos insetos usando a "evo-devo", além de evidências paleontológicas. Isso sugere que as asas são homólogas em série com as estruturas tergal e pleural, possivelmente resolvendo o debate secular.[58] Jakub Prokop e seus colegas encontraram em 2017 evidências paleontológicas de almofadas de asas de ninfas do Paleozoico de que as asas de fato tiveram essa origem dupla.[59]
Ver também
Referências
- ↑ a b c d Chapman, A. D. (2006). Numbers of living species in Australia and the World. [S.l.]: Canberra: Australian Biological Resources Study. pp. 60pp. ISBN 978-0-642-56850-2. Consultado em 24 de junho de 2025. Cópia arquivada em 19 de maio de 2009
- ↑ Smith, D.S. (1965). «Flight muscles of insects». Scientific American. 212 (6): 76–88. Bibcode:1965SciAm.212f..76S. PMID 14327957. doi:10.1038/scientificamerican0665-76
- ↑ Josephson, Robert K.; Malamud, Jean G.; Stokes, Darrell R. (2001). «The efficiency of an asynchronous flight muscle from a beetle». The Journal of Experimental Biology. 204 (23): 4125–4139. PMID 11809787. doi:10.1242/jeb.204.23.4125
- ↑ «Definition of Asynchronous muscle in the Entomologists' glossary». Department of Entomology, North Carolina State University. Consultado em 21 de março de 2011
- ↑ Deora, Tanvi; Gundiah, Namrata; Sane, Sanjay P. (15 de abril de 2017). «Mechanics of the thorax in flies». The Journal of Experimental Biology. 220 (8): 1382–1395. PMID 28424311. doi:10.1242/jeb.128363

- ↑ Heide, G.G. (1983). «Neural mechanisms of flight control in Diptera». In: Nachtigall, Werner. BIONA Report. 2: Insect Flight II. Stuttgart: Gustav Fischer. pp. 33–52
- ↑ Melis, Johan M.; Dickinson, Michael H. (30 de junho de 2023). «Machine learning reveals the control mechanics of the insect wing hinge». BioRxiv: The Preprint Server for Biology. doi:10.1101/2023.06.29.547116
- ↑ a b c d e f g h i j k Wang, Z. Jane. «Dissecting Insect Flight» (PDF). Annual Review of Fluid Mechanics. 37 (1): 183–210. Bibcode:2005AnRFM..37..183W. doi:10.1146/annurev.fluid.36.050802. Consultado em 24 de junho de 2025
- ↑ a b c d e f g Sane, Sanjay P. (2003). «The aerodynamics of insect flight» (PDF). The Journal of Experimental Biology. 206 (23): 4191–4208. PMID 14581590. doi:10.1242/jeb.00663. Consultado em 24 de junho de 2025
- ↑ Wagner, Herbert (1925). «Über die Entstehung des dynamischen Auftriebes von Tragflügeln» [Sobre a origem do levantamento dinâmico de asas]. Zeitschrift für Angewandte Mathematik und Mechanik (em alemão). 5 (1): 17–35. doi:10.1002/zamm.19250050103
- ↑ «Transition and Turbulence». Princeton University. Consultado em 24 de junho de 2025
- ↑ Ellis, D. G.; Stollery, J. L. (1988). «The Behaviour and Performance of Leading-Edge Vortex Flaps» (PDF). International Council of Aeronautical Sciences. ICAS 1988 Jerusalem. 1988: 758–765. Consultado em 24 de junho de 2025
- ↑ Himmelskamp, H. (1945) "Profile investigations on a rotating airscrew". PhD thesis, University of Göttingen.
- ↑ a b c d e f g h i j k l m n o p q r s t u v Davidovits, Paul (2008). Physics in Biology and Medicine. [S.l.]: Academic Press. pp. 78–79. ISBN 978-0-12-369411-9. Consultado em 24 de junho de 2025
- ↑ Hope, Daniel K; DeLuca, Anthony M.; O'Hara, Ryan P (3 de janeiro de 2018). «Investigation into Reynolds number effects on a biomimetic flapping wing». International Journal of Micro Air Vehicles. 10 (1): 106–122. doi:10.1177/1756829317745319
- ↑ «Catching the Wake». Scientific American. Junho de 1999
- ↑ Weis-Fogh, Torkel (1973). «Quick estimates of flight fitness in hovering animals, including novel mechanisms of lift production». Journal of Experimental Biology. 59: 169–230. doi:10.1242/jeb.59.1.169
- ↑ a b Santhanakrishnan, A.; Robinson, A.; Jones, S.; Low, A.; Gadi, S.; Hendrick, T.; Miller, L. (2014). «Clap and fling mechanism with interacting porous wing in tiny insect flight». The Journal of Experimental Biology. 217: 3898–4709. PMID 25189374. doi:10.1242/jeb.084897
- ↑ a b Kolomenskiy, D; Moffatt, H.; Farge, M.; Schneider, K. (2011). «Two- and three- dimensional numerical simulations of the clap-fling-sweep of hovering insects». Journal of Fluids and Structures. 27 (5): 784. Bibcode:2011JFS....27..784K. doi:10.1016/j.jfluidstructs.2011.05.002. Consultado em 24 de junho de 2025
- ↑ Bennett, L. (1977). «Clap and fling aerodynamics- an experimental evaluation». Journal of Experimental Biology. 69: 261–272. doi:10.1242/jeb.69.1.261
- ↑ a b c Miller, L.; Peskin, S. (2009). «Flexible clap and fling in tiny insect flight». The Journal of Experimental Biology. 212 (19): 3076–3090. PMID 19749100. doi:10.1242/jeb.028662
- ↑ a b Lehmann, F.-O.; Sane, S.; Dickinson, M. (2005). «The aerodynamic effects of wing-wing interaction in flapping insect wings». Journal of Experimental Biology. 208 (Pt 16): 3075–3092. PMID 16081606. doi:10.1242/jeb.01744
- ↑ Lehmann, F.-O.; Pick, S. (2007). «The aerodynamic benefit of wing-wing interaction depends on stroke trajectory in flapping insect wings». Journal of Experimental Biology. 210 (Pt 8): 1362–1377. PMID 17401119. doi:10.1242/jeb.02746
- ↑ a b c d Cheng, Xin; Sun, Mao (11 de maio de 2016). «Wing-kinematics measurement and aerodynamics in a small insect in hovering flight». Scientific Reports. 6 (1): 25706. Bibcode:2016NatSR...625706C. PMID 27168523. doi:10.1038/srep25706
- ↑ Kasoju, V.; Santhanakrishnan, A. (2021). «Aerodynamic interaction of bristled wing pairs in fling». Physics of Fluids. 33 (3): 031901. Bibcode:2021PhFl...33c1901K. arXiv:2011.00939. doi:10.1063/5.0036018
- ↑ Weisberger, Mindy (19 de fevereiro de 2016). «Swim Like a Butterfly? Sea Snail 'Flies' Through Water». Scientific American. Consultado em 24 de junho de 2025
- ↑ Murphy, D.; Adhikari, D.; Webster, D.; Yen, J. (2016). «Underwater flight by the planktonic sea butterfly». Journal of Experimental Biology. 219 (4): 535–543. PMID 26889002. doi:10.1242/jeb.129205
- ↑ «Insect Wings in General». Aerodynamics of Insects. Cislunar Aerospace. 1997. Consultado em 28 de março de 2011
- ↑ «Butterflies in the Pieridae family (whites)». Bumblebee.org. Consultado em 18 de março de 2018
- ↑ a b Dudley, Robert (2000). The Biomechanics of Insect Flight: Form, Function, Evolution. Princeton, New Jersey: Princeton University Press. p. 69. JSTOR j.ctv301g2x
- ↑ a b Stocks, Ian. (2008). Sec.Wing Coupling in Capinera (Ed) (2008) "Encyc. Entom.", Vol 4. p. 4266
- ↑ Scoble, Malcolm J. (1995). The Lepidoptera: Form, Function and Diversity. [S.l.]: Natural History Museum. pp. 56–60. ISBN 978-0-19-854952-9. Consultado em 24 de junho de 2025
- ↑ a b Powell, Jerry A. Ch. Lepidoptera (pp. 631-664) in Resh, & Cardé (Eds). "Encyclopedia of Insects". 2003.
- ↑ Gorb, S. (2001) Ch 4.1.5 "Inter-locking of body parts". pp 46–50.
- ↑ Woiwod, I.P.; Reynolds, D.R.; Thomas, C.D. (Eds) 2001. Insect Movement: Mechanisms and Consequences. CAB International.
- ↑ Joos, Barbara (1 de novembro de 1987). «Carbohydrate Use in the Flight Muscles of Manduca Sexta During Pre-Flight Warm-Up». Journal of Experimental Biology (em inglês) (1): 317–327. ISSN 0022-0949. doi:10.1242/jeb.133.1.317. Consultado em 7 de julho de 2025
- ↑ Sane, Sanjay P., Alexandre Dieudonné, Mark A. Willis, and Thomas L. Daniel. "Antennal mechanosensors mediate flight control in moths." science 315, no. 5813 (2007): 863-866.
- ↑ Dickerson, Bradley H., Alysha M. de Souza, Ainul Huda, and Michael H. Dickinson. "Flies regulate wing motion via active control of a dual-function gyroscope." Current Biology 29, no. 20 (2019): 3517-3524.
- ↑ Wolf, Harald. "The locust tegula: significance for flight rhythm generation, wing movement control and aerodynamic force production." Journal of Experimental Biology 182, no. 1 (1993): 229-253.
- ↑ Misof, Bernhard; Liu, Shanlin; Meusemann, Karen; Peters, Ralph S.; Donath, Alexander; Mayer, Christoph; Frandsen, Paul B.; Ware, Jessica; Flouri, Tomáš (7 de novembro de 2014). «Phylogenomics resolves the timing and pattern of insect evolution». Science (em inglês) (6210): 763–767. ISSN 0036-8075. doi:10.1126/science.1257570. Consultado em 7 de julho de 2025
- ↑ Brauckmann, Carsten; Schneider, Jörg (1 de fevereiro de 1996). «A Lower Carboniferous insect from the Bitterfeld/Delitzsch area (Pterygota, Arnsbergian, Germany)». Neues Jahrbuch für Geologie und Paläontologie - Monatshefte (em inglês) (1): 17–30. ISSN 0028-3630. doi:10.1127/njgpm/1996/1996/17. Consultado em 7 de julho de 2025
- ↑ Tillyard, R. J. (agosto de 1928). «SOME REMARKS ON THE DEVONIAN FOSSIL INSECTS FROM THE RHYNIE CHERT BEDS, OLD RED SANDSTONE». Transactions of the Royal Entomological Society of London (em inglês) (1): 65–71. ISSN 0035-8894. doi:10.1111/j.1365-2311.1928.tb01188.x. Consultado em 7 de julho de 2025
- ↑ Haug, Carolin; Haug, Joachim (2017). «The presumed oldest flying insect: more likely a myriapod?». PeerJ. 5: e3402. PMC 5452959. PMID 28584727. doi:10.7717/peerj.3402
- ↑ Wipfler, Benjamin; et al. (19 de fevereiro de 2019). «Evolutionary history of Polyneoptera and its implications for our understanding of early winged insects». Proceedings of the National Academy of Sciences. 116 (8): 3024–3029. Bibcode:2019PNAS..116.3024W. PMC 6386694. PMID 30642969. doi:10.1073/pnas.1817794116
- ↑ Yanoviak, Stephen P.; Dudley, Robert (julho de 2018). «Jumping and the aerial behavior of aquatic mayfly larvae (Myobaetis ellenae, Baetidae)». Arthropod Structure & Development. 47 (4): 370–374. Bibcode:2018ArtSD..47..370Y. PMID 28684306. doi:10.1016/j.asd.2017.06.005
- ↑ a b Trueman, J. W. H. (1990), Comment: evolution of insect wings: a limb exite plus endite model. Canadian Journal of Zoology.
- ↑ a b Crampton, G. (1916). «The Phylogenetic Origin and the Nature of the Wings of Insects According to the Paranotal Theory». Journal of the New York Entomological Society. 24 (1): 1–39. JSTOR 25003692
- ↑ a b c Grimaldi, David; Engel, Michael S. (2005). Insects take to the skies. Evolution of the Insects. New York: Cambridge University Press. pp. 155–159
- ↑ Müller, Fritz (1875) Jena. Zeitung Naturwissenschaften, 9, 241.
- ↑ Kukalova‐Peck, Jarmila (abril de 1978). «Origin and evolution of insect wings and their relation to metamorphosis, as documented by the fossil record». Journal of Morphology (em inglês) (1): 53–125. ISSN 0362-2525. doi:10.1002/jmor.1051560104. Consultado em 7 de julho de 2025
- ↑ Rasnitsyn, Alexander P. (junho de 1981). «A modified paranotal theory of insect wing origin». Journal of Morphology (em inglês) (3): 331–338. ISSN 0362-2525. doi:10.1002/jmor.1051680309. Consultado em 7 de julho de 2025
- ↑ Thomas, Adrian L.R.; Åke Norberg, R. (maio de 1996). «Skimming the surface — the origin of flight in insects?». Trends in Ecology & Evolution (em inglês) (5): 187–188. doi:10.1016/0169-5347(96)30022-0. Consultado em 7 de julho de 2025
- ↑ Marden, James (2003). «The Surface-Skimming Hypothesis for the Evolution of Insect Flight». Acta Zoologica Cracoviensia. 46: 73–84
- ↑ Yanoviak, Stephen P; Kaspari, Michael; Dudley, Robert (23 de agosto de 2009). «Gliding hexapods and the origins of insect aerial behaviour». Biology Letters (em inglês) (4): 510–512. ISSN 1744-9561. PMC 2781901. PMID 19324632. doi:10.1098/rsbl.2009.0029. Consultado em 7 de julho de 2025
- ↑ Averof, Michalis; Cohen, Stephen M. (fevereiro de 1997). «Evolutionary origin of insect wings from ancestral gills». Nature (em inglês) (6617): 627–630. ISSN 0028-0836. doi:10.1038/385627a0. Consultado em 8 de julho de 2025
- ↑ Niwa, Nao; Akimoto‐Kato, Ai; Niimi, Teruyuki; Tojo, Koji; Machida, Ryuichiro; Hayashi, Shigeo (março de 2010). «Evolutionary origin of the insect wing via integration of two developmental modules». Evolution & Development (em inglês) (2): 168–176. ISSN 1520-541X. doi:10.1111/j.1525-142X.2010.00402.x. Consultado em 8 de julho de 2025
- ↑ Elias-Neto, Moysés; Belles, Xavier (agosto de 2016). «Tergal and pleural structures contribute to the formation of ectopic prothoracic wings in cockroaches». Royal Society Open Science (em inglês) (8). 160347 páginas. ISSN 2054-5703. PMC 5108966. PMID 27853616. doi:10.1098/rsos.160347. Consultado em 8 de julho de 2025
- ↑ Tomoyasu, Yoshinori; Ohde, Takahiro; Clark-Hachtel, Courtney (14 de março de 2017). «What serial homologs can tell us about the origin of insect wings». F1000Research (em inglês). 268 páginas. ISSN 2046-1402. PMC 5357031. PMID 28357056. doi:10.12688/f1000research.10285.1. Consultado em 8 de julho de 2025
- ↑ Prokop, Jakub; Pecharová, Martina; Nel, André; Hörnschemeyer, Thomas; Krzemińska, Ewa; Krzemiński, Wiesław; Engel, Michael S. (janeiro de 2017). «Paleozoic Nymphal Wing Pads Support Dual Model of Insect Wing Origins». Current Biology (em inglês) (2): 263–269. doi:10.1016/j.cub.2016.11.021. Consultado em 8 de julho de 2025
Ligações externas
- Animais que voam e planam
- Migração de insetos
- O papel de um inseto no desenvolvimento de microveículos aéreos
- Microveículos aéreos com asas que se movem como insetos
- A nova aerodinâmica do voo dos insetos: Aplicações a Micro Veículos Aéreos
- A aerodinâmica do voo dos insetos
- Visualização de fluxo de mecanismos aerodinâmicos de borboletas
- Voo de insetos