Triaenops goodmani
Triaenops goodmani
| |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Holoceno Inferior | |||||||||||||||||
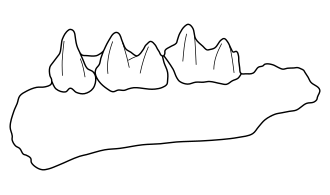 Mandíbula | |||||||||||||||||
| Estado de conservação | |||||||||||||||||
 Extinta (IUCN 3.1) [1] | |||||||||||||||||
| Classificação científica | |||||||||||||||||
| |||||||||||||||||
| Nome binomial | |||||||||||||||||
| †Triaenops goodmani Samonds, 2007 | |||||||||||||||||
| Distribuição geográfica | |||||||||||||||||
![Local de coleta de Triaenops goodmani (em marrom) e distribuição das espécies vivas de Triaenops (verde -T. menamena; azul -T. afer; vermelho -T. persicus; amarelo -T. persicus e T. parvus.[2]](./_assets_/0c70a452f799bfe840676ee341124611/Triaenops_distribution.png) Local de coleta de Triaenops goodmani (em marrom) e distribuição das espécies vivas de Triaenops (verde -T. menamena; azul -T. afer; vermelho -T. persicus; amarelo -T. persicus e T. parvus.[2]
| |||||||||||||||||
Triaenops goodmani é um morcego extinto de Madagascar pertencente ao gênero Triaenops.[1] Conhecido a partir de três mandíbulas inferiores coletadas em uma caverna em Anjohibe em 1996, foi descrito como uma nova espécie em 2007. O material tem, no máximo, 10.000 anos de idade. Um úmero (osso do braço superior) de morcego encontrado no mesmo local não pôde ser identificado como pertencente a T. goodmani ou à espécie viva T. menamena. T. goodmani é identificado como membro do gênero Triaenops ou do gênero relacionado Paratriaenops por características dentárias, como o quarto pré-molar semelhante a um canino com uma única cúspide e a presença de um espaço entre as cúspides entoconídio e hipoconulídio nos dois primeiros molares. T. goodmani é maior que as espécies vivas de Triaenops e Paratriaenops em Madagascar, e no primeiro molar, a cúspide protoconídio é apenas ligeiramente mais alta que o hipoconídio, diferentemente das outras espécies, onde o protoconídio é significativamente mais alto.
Taxonomia e distribuição
Em 1996, uma equipe liderada por David Burney coletou brechas contendo restos de morcegos e outros animais na caverna de Anjohibe, no noroeste de Madagascar.[3] Os morcegos da amostra foram descritos por Karen Samonds (anteriormente Irwin) em sua dissertação de doutorado de 2006 e em um artigo de 2007.[4] Ela identificou várias espécies vivas, além de duas espécies extintas descritas como novas, Triaenops goodmani e Hipposideros besaoka.[5] Na época, o gênero Triaenops era considerado como incluindo três espécies em Madagascar: Triaenops auritus, Triaenops furculus e Triaenops rufus.[6] Posteriormente, Steven M. Goodman [en] e Julie Ranivo descobriram que o nome rufus não se aplicava às espécies de Madagascar, propondo o nome Triaenops menamena para os morcegos malgaxes anteriormente conhecidos como Triaenops rufus.[7]
Além disso, Petr Benda e Peter Vallo transferiram as outras duas espécies malgaxes para um gênero separado, Paratriaenops, agora conhecidas como Paratriaenops auritus e Paratriaenops furculus.[8] O nome específico da espécie extinta, goodmani, homenageia Steven M. Goodman por suas pesquisas sobre morcegos malgaxes.[6] O material de T. goodmani é proveniente da localidade OLD SE dentro da caverna e tem cerca de 10.000 anos ou menos.[9] Uma análise cladística usando dados de morfologia não conseguiu resolver as relações de Triaenops goodmani, mas não o colocou junto às outras espécies de Triaenops e Paratriaenops estudadas.[10] Em um artigo de 2008, Amy Russell e colegas comentaram que características cranianas de T. goodmani sugerem que ele pertence ao grupo "T. furculus/T. auritus", agora classificado em Paratriaenops.[11]
Descrição
Triaenops goodmani é conhecido por três mandíbulas inferiores: uma com o quarto pré-molar (p4) e os primeiro e segundo molares (m1–2), e duas com o segundo e terceiro molares (m2–3).[6] A mandíbula é relativamente robusta.[12] O p4 assemelha-se a um canino, com uma única cúspide aproximadamente tão alta quanto a cúspide mais alta do m1, sem plataformas ou cúspides acessórias. Os molares têm coroas estreitas e são mais longos do que em T. menamena, P. auritus e P. furcula.[6] O comprimento do m2 varia de 1,55 a 1,57 mm, e a largura de 0,98 a 1,02 mm.[13] No m1, o trigonídio (grupo frontal de cúspides) é mais estreito e ligeiramente mais alto que o talonídio na parte posterior. O protoconídio, uma das principais cúspides do trigonídio, é a mais alta,[14] mas apenas ligeiramente mais alta que o hipoconídio (uma cúspide do talonídio); nas espécies vivas de Triaenops e Paratriaenops de Madagascar, o protoconídio é significativamente mais alto que o hipoconídio. O paraconídio, metaconídio (ambos cúspides do trigonídio) e entoconídio (uma cúspide do talonídio) são mais baixos do que em Paratriaenops auritus.[6] O hipoconulídio (parte do talonídio) é pequeno, mas distinto, sendo a cúspide mais baixa, separada do entoconídio por um espaço.[14] T. goodmani não possui uma crista conectando o entoconídio ao metaconídio.[15] Há uma crista (cíngulo) na parte frontal e posterior do dente.[6] Os dois últimos molares são semelhantes ao m1, mas no m2 o talonídio é apenas ligeiramente mais largo que o trigonídio, e no m3 ambos têm a mesma largura.[14] Além disso, há uma plataforma entre o protoconídio e o hipoconídio no m2,[6] e o m3 é menor, não apresenta o espaço entre o entoconídio e o hipoconulídio, e tem uma crista fraca entre o entoconídio e o metaconídio.[14] Essas características são típicas de Triaenops e Paratriaenops.[12]
No mesmo local onde T. goodmani foi encontrado, Samonds também registrou a extremidade distal (mais afastada) de um úmero de Triaenops, com uma largura de 3,58 mm. Esse osso era semelhante aos úmeros de T. menamena, mas ela não o identificou como pertencente a nenhuma das espécies devido à pequena diferença de tamanho entre T. menamena e T. goodmani.[16] No sítio NCC-1 (estimado entre 69.600 e 86.800 anos de idade),[17] foram registradas duas mandíbulas de Triaenops, uma com p4 e m1, e outra com m1–2 e parte do m3.[18] Em relação às espécies vivas de Triaenops e Paratriaenops, o m1 dessas mandíbulas é mais longo e estreito. Embora as amostras sejam pequenas, as medidas não correspondem às de T. goodmani. Além disso, a crista entre o entoconídio e o metaconídio é mais forte do que em T. goodmani. Samonds identificou essas mandíbulas apenas como Triaenops.[19]
Referências
- ↑ a b Samonds, 2007, p. 62
- ↑ Samonds, 2007, fig. 1; Benda e Vallo, 2009, fig. 1
- ↑ Samonds, 2007, pp. 40–41
- ↑ Samonds, 2006; 2007
- ↑ Samonds, 2007, p. 39
- ↑ a b c d e f g Samonds, 2007, p. 46
- ↑ Goodman e Ranivo, 2009
- ↑ Benda e Vallo, 2009, p. 34
- ↑ Samonds, 2007, pp. 42–43
- ↑ Samonds, 2006, p. 178, figs. 4.6, 4.7
- ↑ Russell et al., 2008, p. 1001
- ↑ a b Samonds, 2007, p. 48
- ↑ Samonds, 2007, tabela 3
- ↑ a b c d Samonds, 2007, p. 47
- ↑ Samonds, 2007, pp. 46–47
- ↑ Samonds, 2007, p. 49
- ↑ Samonds, 2007, p. 43
- ↑ Samonds, 2007, p. 55
- ↑ Samonds, 2007, p. 57
Bibliografia
- Benda, P. e Vallo, P. 2009. Taxonomic revision of the genus Triaenops (Chiroptera: Hipposideridae) with description of a new species from southern Arabia and definitions of a new genus and tribe. Folia Zoologica 58(Monograph 1):1–45.
- Goodman, S.M. e Ranivo, J. 2009. The geographical origin of the type specimens of Triaenops rufus and T. humbloti (Chiroptera: Hipposideridae) reputed to be from Madagascar and the description of a replacement species name. Mammalia 73:47–55.
- Russell, A.L., Goodman, S.M. e Cox, M.P. 2008. Coalescent analyses support multiple mainland-to-island dispersals in the evolution of Malagasy Triaenops bats (Chiroptera: Hipposideridae). Journal of Biogeography 35:995–1003.
- Samonds (Irwin), K.E. 2006. The origin and evolution of Malagasy bats: Implications of new Late Pleistocene fossils and cladistic analyses for reconstructing biogeographic history. Ph.D. Dissertation, Department of Anatomical Sciences, Stony Brook University, xx + 403 pp.
- Samonds, K.E. 2007. Late Pleistocene bat fossils from Anjohibe Cave, northwestern Madagascar. Acta Chiropterologica 9(1):39–65.