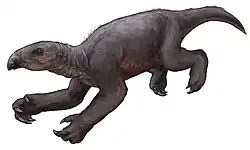
Thalassocnus
Thalassocnus
| |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Mioceno Superior – Plioceno Superior (Huayqueriano [en]–Uquiano [en]) ~7–3 Ma | |||||||||||||||||
| Classificação científica | |||||||||||||||||
| |||||||||||||||||
| Espécie-tipo | |||||||||||||||||
| †Thalassocnus natans de Muizon & McDonald, 1995 | |||||||||||||||||
| Outras espécies | |||||||||||||||||
| |||||||||||||||||

Thalassocnus é um gênero extinto de preguiça-gigante semiaquática do Mioceno e Plioceno (7–3 Ma) da costa pacífica da América do Sul. É monotípico, sendo o único gênero da subfamília Thalassocninae. As cinco espécies — T. antiquus, T. natans, T. littoralis, T. carolomartini e T. yaucensis — representam uma cronoespécie, uma população que se adaptou gradualmente à vida marinha em uma linhagem direta. São as únicas preguiças aquáticas conhecidas, embora também possam ter sido adaptadas à vida terrestre. Fósseis foram encontrados na formação Pisco do Peru, na formação Tafna da Argentina,[2] e nas formações Bahía Inglesa, Coquimbo e Horcón do Chile. A subfamília Thalassocninae foi classificada anteriormente tanto na família Megatheriidae[3] quanto em Nothrotheriidae.[4]
Thalassocnus desenvolveu adaptações à vida marinha ao longo de 4 milhões de anos, como ossos densos e pesados para contrabalançar a flutuação, narinas internas migrando para dentro da cabeça para facilitar a respiração submersa, focinho alargado e alongado para consumir plantas aquáticas, e cabeça inclinada para baixo, auxiliando na alimentação no fundo do mar. A cauda longa era provavelmente usada para mergulho e equilíbrio, semelhante ao castor e ao ornitorrinco.
Thalassocnus caminhava pelo fundo do mar e desenterrava alimentos com suas garras. Não realizava natação potente, recorrendo à remada quando necessário. Espécies iniciais eram provavelmente generalistas, consumindo macroalgas e ervas-marinhas perto da costa, enquanto espécies posteriores se especializaram em ervas-marinhas mais afastadas. Eram possivelmente presas de tubarões e cachalotes como Acrophyseter. Fósseis foram encontrados em formações com grandes assembleias de mamíferos marinhos e tubarões.
Etimologia
O nome genérico Thalassocnus deriva das palavras em grego thalassa ("mar") e Ocnus, uma divindade alegórica da mitologia grega e romana que representa a perda de tempo ou preguiça.
O nome da espécie carolomartini homenageia Carlos Martin, falecido proprietário da fazenda Sacaco e descobridor de vários ossos na formação Pisco, incluindo o espécime holótipo;[5] e yaucensis refere-se ao vilarejo Yauca, próximo ao local de descoberta.[6]
Taxonomia
Espécimes-tipo

Thalassocnus viveu durante o período Mioceno Superior ao final do Plioceno e todas as cinco espécies foram descobertas em diferentes horizontes da formação Pisco no Peru. T. antiquus foi encontrado no horizonte Aguada de Lomas, em estratos geológicos de 7 ou 8 milhões de anos; T. natans (espécie-tipo), no horizonte Montemar, viveu há cerca de 6 milhões de anos; T. littoralis, no horizonte Sud-Sacaco, há cerca de 5 milhões de anos; T. carolomartini, no horizonte Sacaco, entre 3 e 4 milhões de anos; e T. yaucensis, no horizonte Yuaca, de 3 a 1,5 milhões de anos.[6] Espécimes também foram encontrados na formação Bahía Inglesa,[7] na formação Coquimbo e na formação Horcón no Chile.[8] Três espécies foram confirmadas nas formações chilenas: T. carolomartini, T. natans e T. antiquus, com a presença de T. yaucensis considerada provável.[9]
Em 1995, o gênero Thalassocnus foi formalmente descrito com a espécie T. natans, com base em um esqueleto parcial pelos paleontólogos Christian de Muizon e H. Gregory McDonald.[10] Em 2002, T. littoralis foi descrito a partir de um crânio quase completo e T. carolomartini foi descrito com base em um crânio e mãos, possivelmente do mesmo indivíduo.[5] T. antiquus foi descrito em 2003, incluindo crânio, mandíbula e grande parte do corpo, embora danificado.[11] T. yaucensis foi descrito em 2004 a partir de um crânio e esqueleto quase completo.[6]
Filogenia

Em 1968, o taxonomista Robert Hoffstetter classificou restos de preguiça não descritos na família Megatheriidae, possivelmente na subfamília extinta Planopsinae, com base em semelhanças com o osso do tornozelo e fêmur.[12] Em 1995, foram movidos para a subfamília Nothrotheriinae.[10] Em 2004, esta foi elevada à família Nothrotheriidae, e as preguiças foram colocadas na nova subfamília Thalassocninae.[6] Em 2017, foram reclassificados para a família Megatheriidae, com a subfamília Thalassocninae possivelmente divergindo de Megatheriinae durante o período Friasiano do Mioceno, há cerca de 16 milhões de anos.[3] Uma análise de 2018 mantém a subfamília Thalassocninae na família Nothrotheriidae.[4] Como as duas famílias podem ser irmãs,[13] e a posição da subfamília Thalassocninae seria provavelmente basal, a classificação familiar correta é desafiadora.
As cinco espécies formam uma linhagem direta (cronoespécie), mas é possível que T. antiquus não seja ancestral de T. natans.[14][11][10]
| |||||||||||||||||||||||||||||||||||||||||||||
| Filogenia da subfamília Thalassocninae, assumindo colocação na família Megatheriidae[3] |
Descrição
Tamanho
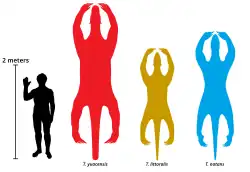
Thalassocnus é o único integrante aquático dos Xenarthra, grupo que inclui preguiças, tamanduás e tatus, embora as preguiças-gigantes Eionaletherium do Mioceno da Venezuela e Ahytherium do Pleistoceno do Brasil possam ter se adaptado à vida litorânea.[15] Contudo, Thalassocnus também era adaptado à vida terrestre, com base em registros na Argentina.[2] Seu tamanho aumentou ao longo do tempo.[5]
T. natans tem o esqueleto mais completo, medindo 2,55 m do focinho à cauda. Com base na proporção entre o fêmur e o corpo, o espécime de T. littoralis, provavelmente fêmea, media 2,1 m, e o de T. yaucensis, 3,3 m.[16]
Crânio

Espécies posteriores tinham pré-maxilas alargadas, resultando em focinho alongado. A mandíbula inferior se alongou progressivamente e passou a apresentar forma de colher, imitando a função dos incisivos abertos de ruminantes. Possuíam lábios fortes, indicados pelo grande forame infraorbital. O focinho era quadrado, diferente do triangular de folívoros. As narinas migraram para o topo do focinho, como em focas.[16] Para alimentação submersa, o palato mole separando traqueia e esôfago era desenvolvido, e as narinas internas ficavam mais internas, ampliando a boca. Essas adaptações, também vistas em mamíferos terrestres, podem estar ligadas à eficiência de mastigação. O músculo masseter era o principal para mordida, mais potente em espécies posteriores para agarrar ervas-marinha. O músculo pterigóideo lateral era maior, adaptado para triturar.[14][5] As espécies T. carolomartini e T. yaucensis sugerem uma tromba curta, como em antas e elefantes-marinhos.[17]
O Thalassocnus tinha dentição hipsodonte com coroa dentária alta e esmalte dentário além da gengiva. Não possuía caninos, com quatro molares superiores e três inferiores de cada lado. Os dentes tinham durodentina externa, um tipo de dentina similar a ossos, e vasodentina interna, uma dentina que permite circulação de sangue. Possuíam formato de prisma, com seção transversal circular, interligando-se na mastigação em espécies posteriores. Espécies iniciais afiavam os dentes através da mastigação; tinham dentes retangulares, como a preguiça-gigante Megatherium americanum. Espécies posteriores tinham dentes quadrados e maiores, para triturar.[14][5]
Vértebras
Thalassocnus possuía 7 vértebras cervicais; 17 vértebras torácicas, enquanto os membros da família Megatheriidae apresentavam 18; 3 vértebras lombares; e 24 vértebras caudais, diferente de outras preguiças, que possuem menos de 20. Os centros vertebrais encurtaram, estabilizando a coluna, possibilitando uma adaptação para escavação.[16]
Em espécies posteriores, os processos espinhosos das vértebras torácicas eram mais altos que os cervicais, diferente de outras preguiças. Vértebras cervicais pequenas indicam músculos do pescoço fracos, com a cabeça inclinada para baixo em repouso. A articulação entre a primeira vertebra e o crânio era forte, adaptação para alimentação no fundo do mar.[16] A coluna tornou-se rígida, e a cabeça alinhava-se com a espinha, como em baleias.[5]
O processo espinhoso da primeira vértebra torácica é quase vertical, mas as demais inclinam-se para a cauda, com inclinação maior em espécies posteriores: 70° em T. littoralis e T. carolomartini, diferente da inclinação de 30° em T. antiquus e T. natans. A inclinação diminui na nona vértebra torácica, sugerindo músculos dorsais menos desenvolvidos para natação.[16]
As vértebras caudais indicam musculatura forte, semelhante ao castor (Castor spp.) e ornitorrinco (Ornithorhynchus anatinus), usando a cauda para equilíbrio e mergulho. A cauda longa era adaptação para mergulho, como em corvos-marinhos (Phalacrocorax spp.), gerando sustentação descendente.[16]
Membros
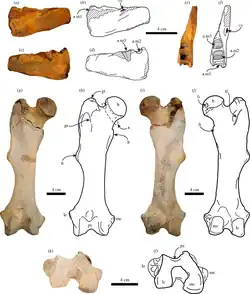

Espécies posteriores tinham músculos de braço fortes, indicados por grandes fossas nas escápulas, cotovelo e punho, e braços curtos, para escavação. Possuíam cinco garras.[16]
Pernas mais estreitas e cristas ilíacas reduzidas indicam menor dependência delas, contando com flutuação, e repouso semi-submerso. As pernas tornaram-se flexíveis, com fêmures alcançando posição horizontal em espécies posteriores. Fêmur e patela são menores em T. yaucensis que em T. natans. Diferente de preguiças-gigantes, que sobrecarregam membros traseiros no bipedismo, os ossos de Thalassocnus são esguios, com tíbia e fêmur de comprimentos similares.[18]
Espécies iniciais apoiavam-se nas laterais dos pés (pedolateral); posteriores eram plantígrados, para remar e caminhar no fundo do mar. Dígitos reduziram-se, com o terceiro dígito flexionado para ancorar no fundo. O quinto dígito era vestigial.[18]
Paleobiologia

Densidade óssea
Ossos densos (paquiosteosclerose) em espécies posteriores combatiam a flutuação, permitindo afundar, como em peixes-bois. T. antiquus tinha densidade similar a preguiças-gigantes. Em espécies posteriores, a cavidade medular era quase ausente nos membros, que pesavam mais no esqueleto. Essa condição, vista em baleias do grupo Archaeoceti, desenvolveu-se em 4 milhões de anos. Preguiças da ordem Pilosa têm ossos densos, predispondo à exaptação.[16][19]
Metabolismo
Valores de δ18Op de ossos e dentes indicam taxa metabólica elevada, herdada de ancestrais terrestres.[20]
Forrageamento

Herbívoros costeiros, tornaram-se aquáticos devido à desertificação e escassez de alimentos terrestres.[5] Espécies iniciais eram generalistas, forrageando algas e ervas marinhas em costas arenosas, com marcas de areia nos dentes, em profundidades menores que um metro. T. antiquus consumia plantas levadas à costa. Espécies posteriores, T. carolomartini e T. yaucensis, alimentavam-se em águas profundas, como peixes-bois. Espécies iniciais mastigavam verticalmente; posteriores, anteroposteriormente.[14][16]
Espécies posteriores alimentavam-se no fundo, como sirenídeos e desmostílios, especializando-se em ervas-marinhas, como espécies da família Zosteraceae da formação Pisco, incluindo a enguia-marinha Zostera tasmanica.[14][21] Caminhavam no fundo do mar, sem adaptações para natação potente. Desenterravam ervas-marinhas com garras, como castores e ornitorrincos, e usavam lábios fortes para arrancá-las. A dieta incluía alimentos enterrados.[14][16][18]

Usavam garras para soltar terra, cortar vegetação, agarrar alimentos ou ancorar no fundo do mar. Seguravam rochas em ondas fortes. Tíbia e fíbula quebradas e cicatrizadas sugerem indivíduos lançados contra rochas em tempestades, usando garras para retornar à costa.[14][16]
Competiam com dugongos por ervas-marinhas, embora estes fossem raros.[5] Na formação Bahía Inglesa, em Cerro Ballena, podem ter morrido por proliferação de algas nocivas.[22] Eram predados por Acrophyseter,[23] com indivíduos feridos vulneráveis a tubarões.[14]
Variação sexual
Thalassocnus pode ter exibido dimorfismo sexual, sugerido por variações em fósseis de T. littoralis e entre crânios de T. carolomartini. Diferenças incluem tamanho, esguiez dos dentes e comprimento das pré-maxilas, reminiscentes de lábios superiores ou probóscide em machos de elefante-marinho (Mirounga spp.).[14]
Paleoecologia

Fósseis de Thalassocnus foram encontrados no Peru e Chile, área desértica desde o Mioceno Médio.[7][5]
A formação Pisco é conhecida por vertebrados marinhos, incluindo misticetos da família Cetotheriidae, balenopterídeos Balaenoptera siberi, zifiídeos Messapicetus gregarius, pontoporiídeos Brachydelphis mazeasi, golfinhos da família Delphinidae, cetáceos Odobenocetops e cachalotes Acrophyseter e Livyatan.[24][25] Outros vertebrados incluem focas Acrophoca e Hadrokirus, tartarugas Pacifichelys urbinai, gavialídeos Piscogavialis, corvos-marinhos, procelariídeos e aves do gênero Sula. A formação possui peixes cartilaginosos, como tubarões da ordem Carcharhiniformes e das famílias Carcharhinidae e Sphyrnidae; e, em menor extensão, tubarões da ordem Lamniformes, como espécies das famílias Lamnidae, Odontaspididae e Otodontidae. Muito dentes encontrados foram associados ao megalodonte (Carcharocles megalodon). Outros peixes cartilaginosos incluem espécies das famílias Myliobatidae, Pristidae e Squatinidae. Os peixes ósseos encontrados são atuns e cienídeos. Espécies de Livyatan e o megalodonte provavelmente eram superpredadores.[24][26]
A formação Bahía Inglesa, na bacia de Caldera, tem grande população de tubarões, como megalodonte e tubarão-branco (Carcharodon carcharias). Outros incluem tubarão-azul (Prionace glauca), Odontaspis ferox, Pristiophorus da ordem Pristiophoriformes, tubarões do gênero Squatina, tubarões Heterodontiformes (Heterodontus spp.), arraias do gênero Myliobatis e peixes do gênero Callorhinchus.[7][27] Aves do gênero Pachyptila foram encontradas.[7][28] Mamíferos marinhos incluem balenopterídeos, Acrophoca, cachalote do gênero Scaldicetus e Odobenocetops.[22]
As formações Coquimbo, na baía de Tongoy, e Horcón, na bacia de Valparaíso, apresentam tubarões como megalodonte, tubarão-branco, Carcharodon plicatilis, Isurus oxyrinchus, peixes do gênero Pristiophorus, Odontaspis ferox, tubarões Carcharhinus, Galeorhinus galeus, tubarão-cobre (Carcharhinus brachyurus) e tubarão-albafar (Hexanchus griseus).[29][30]
Pinguins, como espécies do gênero Spheniscus, são conhecidos nas três formações.[31]
Extinção
Thalassocnus foi extinto no final do Plioceno devido ao resfriamento após o fechamento do canal da América Central, que eliminou ervas-marinhas da costa pacífica sul-americana.[5][16] Especialistas em ervas-marinhas, as espécies posteriores desenvolveram flutuação negativa para alimentação no fundo do mar, como dugongos. Isso envolvia caminhada no fundo do mar, como hipopótamos e desmostílios, e escavação. A flutuação negativa exige ossos densos e pouca gordura, dificultando a termorregulação em águas frias, especialmente com o baixo metabolismo basal dos xenartros. Assim, Thalassocnus estava mal adaptado às mudanças, mesmo com ervas-marinhas remanescentes.[16]
Ver também
Referências
- ↑ Thalassocnus. Fossilworks. Retrieved 25 August 2018
- ↑ a b Quiñones, Sofía I.; Zurita, Alfredo E.; Miño-Boilini, Ángel R.; Candela, Adriana M.; Luna, Carlos A. (15 de setembro de 2022). «Unexpected record of the aquatic sloth Thalassocnus (Mammalia, Xenarthra, Folivora) in the upper Neogene of the Puna (Jujuy, Argentina)». Journal of Vertebrate Paleontology. 42 (1): e2109973. Bibcode:2022JVPal..42E9973Q. ISSN 0272-4634. doi:10.1080/02724634.2022.2109973
- ↑ a b c Amson, E.; de Muizon, C.; Gaudin, T. J. (2017). «A reappraisal of the phylogeny of the Megatheria (Mammalia: Tardigrada), with an emphasis on the relationships of the Thalassocninae, the marine sloths». Zoological Journal of the Linnean Society. 179 (1): 217–236. doi:10.1111/zoj.12450

- ↑ a b Varela, L.; Tambusso, P.S.; McDonald, H.G.; Fariña, R.A.; Fieldman, M. (2019). «Phylogeny, Macroevolutionary Trends and Historical Biogeography of Sloths: Insights From a Bayesian Morphological Clock Analysis». Systematic Biology. 68 (2): 204–218. PMID 30239971. doi:10.1093/sysbio/syy058
- ↑ a b c d e f g h i j McDonald, H. G.; de Muizon, C. (2002). «The cranial anatomy of Thalassocnus (Xenarthra, Mammalia), a derived nothrothere from the Neogene of the Pisco Formation (Peru)». Journal of Vertebrate Paleontology. 22 (2): 349–365. doi:10.1671/0272-4634(2002)022[0349:TCAOTX]2.0.CO;2
- ↑ a b c d de Muizon, C.; McDonald, H. G.; Salas-Gismondi, R.; Schmitt, M. U. (2004). «The youngest species of the aquatic sloth Thalassocnus and a reassessment of the relationships of the nothrothere sloths (Mammalia: Xenarthra)». Journal of Vertebrate Paleontology. 24 (2): 387–397. Bibcode:2004JVPal..24..387D. doi:10.1671/2429a. Cópia arquivada em 12 de outubro de 2018
- ↑ a b c d Canto, J.; Salas-Gismondi, R.; Cozzuol, M.; Yáñez, J. (2008). «The aquatic sloth Thalassocnus (Mammalia, Xenarthra) from the late Miocene of North-Central Chile: biogeographic and ecological implications». Journal of Vertebrate Paleontology. 28 (3): 918–922. ISSN 0272-4634. doi:10.1671/0272-4634(2008)28[918:TASTMX]2.0.CO;2
- ↑ De los Arcos, S.; Partarrieu, D.; Carrillo-Briceño, J.; Amson, E. (2017). «The southernmost occurrence of the aquatic sloth Thalassocnus (Mammalia, Tardigrada) in two new Pliocene localities in Chile». Ameghiniana. 54 (4): 351–369. doi:10.5710/AMGH.29.12.2016.3004
- ↑ Peralta-Prato, Javiera; Solórzano, Andrés (2019). «How many species of the aquatic sloth Thalassocnus (Xenarthra: Megatheriidae) were in Chile?: new evidences from the Bahía Inglesa Formation, with a reappraisal of their biochronological affinities». Andean Geology. 46 (3): 693. doi:10.5027/andgeoV46n3-3221
- ↑ a b c de Muizon, C.; McDonald, H. G. (1995). «An aquatic sloth from the Pliocene of Peru». Nature. 375 (6528): 224–227. Bibcode:1995Natur.375..224M. doi:10.1038/375224a0
- ↑ a b de Muizon, C.; McDonald, H. G.; Salas, R.; Urbina, M. (2003). «A new early species of the aquatic sloth Thalassocnus (Mammalia, Xenarthra) from the late Miocene of Peru». Journal of Vertebrate Paleontology. 23 (4): 886–894. Bibcode:2003JVPal..23..886D. doi:10.1671/2361-13
- ↑ Hoffstetter, R. (1968). «Un gisement de vertébrés tertiaires à Sacaco (Sud-Pérou), témoin néogène d'une migration de faunes australes au long de la côte occidentale sudaméricaine» [Um depósito de vertebrados terciários em Sacaco (Sul do Peru), testemunha neogênica de uma migração de faunas austrais ao longo da costa ocidental sul-americana]. Comptes Rendus Hebdomadaires des Séances de l'Académie des Sciences (em francês). 267: 1273–1276
- ↑ Presslee, S.; Slater, G.J.; Pujos, F.; Forasiepi, A.M.; Fischer, R.; Molloy, K.; Mackie, M.; Olsen, J.V.; Kramarz, A.; Taglioretti, M.; Scaglia, F. (julho de 2019). «Palaeoproteomics resolves sloth relationships». Nature Ecology & Evolution. 3 (7): 1121–1130. Bibcode:2019NatEE...3.1121P. PMID 31171860. doi:10.1038/s41559-019-0909-z
- ↑ a b c d e f g h i de Muizon, C.; McDonald, H. G.; Salas, R.; Urbina, M. (2004). «The evolution of feeding adaptations of the aquatic sloth Thalassocnus». Journal of Vertebrate Paleontology. 24 (2): 398–410. Bibcode:2004JVPal..24..398D. JSTOR 4524727. doi:10.1671/2429b
- ↑ Rincón, A. D.; McDonald, H. G.; Solórzano, A.; Flores, M. N.; Ruiz-Ramoni, D. (2015). «A new enigmatic Late Miocene mylodontoid sloth from northern South America». Royal Society Open Science. 2 (2): 140256. Bibcode:2015RSOS....240256R. PMC 4448802. PMID 26064594. doi:10.1098/rsos.140256
- ↑ a b c d e f g h i j k l m Amson, E.; Argot, C.; McDonald, H. G.; de Muizon, C. (2015). «Osteology and functional morphology of the axial postcranium of the marine sloth Thalassocnus (Mammalia, Tardigrada) with paleobiological implications». Journal of Mammalian Evolution. 22 (4): 473–518. doi:10.1007/s10914-014-9280-7
- ↑ De Muizon, Christian; McDonald, H. Gregory; Salas, Rodolfo; Urbina, Mario (2004). «The Youngest Species of the Aquatic Sloth Thalassocnus and a Reassessment of the Relationships of the Nothrothere Sloths (Mammalia: Xenarthra)». Journal of Vertebrate Paleontology. 24 (2): 387–397. Bibcode:2004JVPal..24..387D. JSTOR 4524726. doi:10.1671/2429a
- ↑ a b c Amson, E.; Argot, C.; McDonald, H. G.; de Muizon, C. (2004). «Osteology and functional morphology of the hind limb of the marine sloth Thalassocnus (Mammalia, Tardigrada)» (PDF). Journal of Mammalian Evolution. 22 (3): 355–419. doi:10.1007/s10914-014-9274-5. Cópia arquivada (PDF) em 22 de julho de 2018
- ↑ Amson, E.; de Muizon, C.; Laurin, M.; Argot, C.; Buffrénil, V. de (2014). «Gradual adaptation of bone structure to aquatic lifestyle in extinct sloths from Peru». Proceedings of the Royal Society B: Biological Sciences. 281 (1782). 20140192 páginas. PMC 3973278. PMID 24621950. doi:10.1098/rspb.2014.0192
- ↑ Amiot, Romain; Göhlich, Ursula B.; Lécuyer, Christophe; de Muizon, Christian; Cappetta, Henri; Fourel, François; Héran, Marie-Anne; Martineau, François (7 de julho de 2008). «Oxygen isotope compositions of phosphate from Middle Miocene–Early Pliocene marine vertebrates of Peru». Palaeogeography, Palaeoclimatology, Palaeoecology (em inglês). 264 (1–2): 85–92. Bibcode:2008PPP...264...85A. doi:10.1016/j.palaeo.2008.04.001. Consultado em 21 de novembro de 2024 – via Elsevier Science Direct
- ↑ Kuo, J. (2005). «A revision of the genus Heterozostera (Zosteraceae)». Aquatic Botany. 81 (2): 97–140. Bibcode:2005AqBot..81...97K. doi:10.1016/j.aquabot.2004.10.005
- ↑ a b Pyenson, N. D.; Gutstein, C. S.; Parham, J. F.; Le Roux, J. P.; Chavarría, C. C.; Little, H.; Metallo, A.; Rossi, V.; Valenzuela-Toro, A. M.; Velez-Juarbe, J.; Santelli, C. M.; Rogers, D. R.; Cozzuol, M. A.; Suárez, M. E. (2014). «Repeated mass strandings of Miocene marine mammals from Atacama Region of Chile point to sudden death at sea». Proceedings of the Royal Society B. 281 (1781): 1–9. PMC 3953850. PMID 24573855. doi:10.1098/rspb.2013.3316
- ↑ Lambert, O.; Bianucci, G.; de Muizon, C. (2017). «Macroraptorial sperm whales (Cetacea, Odontoceti, Physeteroidea) from the Miocene of Peru». Zoological Journal of the Linnean Society. 179: 404–474. doi:10.1111/zoj.12456. hdl:11568/814760. Arquivado do original em 22 de julho de 2018
- ↑ a b Parham, J. F.; Pyenson, N. D. (2010). «New sea turtle from the Miocene of Peru and iterative evolution of feeding ecomorphologies since the Cretaceous» (PDF). Journal of Paleontology. 84 (2): 231–247. Bibcode:2010JPal...84..231P. doi:10.1666/09-077R.1. Cópia arquivada (PDF) em 1 de dezembro de 2017
- ↑ Brand, L.; Urbina, M.; Chadwick, A.; DeVries, T. J.; Esperante, R. (2011). «A high resolution stratigraphic framework for the remarkable fossil cetacean assemblage of the Miocene/Pliocene Pisco Formation, Peru». Journal of South American Earth Sciences. 31 (4): 414–425. Bibcode:2011JSAES..31..414B. doi:10.1016/j.jsames.2011.02.015
- ↑ Bianucci, G.; Di Celma, C.; Landini, W.; Post, K.; Tinelli, C.; de Muizon, C.; Gariboldi, K.; Malinverno, E.; Cantalamessa, G.; Gioncada, A.; Collareta, A.; Gismondi, R.; Varas-Malca, R.; Urbina, M.; Lambert, O. (2015). «Distribution of fossil marine vertebrates in Cerro Colorado, the type locality of the giant raptorial sperm whale Livyatan melvillei (Miocene, Pisco Formation, Peru)» (PDF). Journal of Maps. 12 (3). 543 páginas. doi:10.1080/17445647.2015.1048315. hdl:10.1080/17445647.2015.1048315
- ↑ Suárez, M. E.; Encinas, A.; Ward, D. (2006). «An Early Miocene elasmobranch fauna from the Navidad Formation, Central Chile, South America». Cainozoic Research. 4 (1–2): 3–18. Cópia arquivada em 22 de agosto de 2018
- ↑ Sallaberry, M.; Rubilar-Rogers, D.; Suárez, M. E.; Gutstein, C. S. (2007). «The skull of a fossil Prion (Aves: Procellariiformes) from the Neogene (Late Miocene) of northern Chile». Revista Geológica de Chile. 34 (1): 147–154. doi:10.4067/S0716-02082007000100008
- ↑ Staig, F.; Hernández, S.; López, P.; Villafaña, J. A.; Varas, C.; Soto, L. P.; Carrillo-Briceño, J. D. (2015). «Late Neogene elasmobranch fauna from the Coquimbo Formation, Chile». Revista Brasileira de Paleontologia. 18 (2): 261–272. doi:10.4072/rbp.2015.2.07
- ↑ Carrillo-Briceño, J. D.; Gonzales-Barba, G.; Landaeta, M. F.; Nielsen, S. N. (2013). «Condrictios fósiles del Plioceno Superior de la Formación Horcón, Región de Valparaíso, Chile central» [Chondrichthyes fósseis do Plioceno Superior da Formação Horcón] (PDF). Revista Chilena de Historia Natural (em espanhol). 86 (2): 191–206. Bibcode:2013RvCHN..86..191C. doi:10.4067/S0716-078X2013000200008. Cópia arquivada (PDF) em 21 de maio de 2018
- ↑ Hoffmeister, M. C.; Carrillo-Briceño, J. D.; Nielsen, S. N. (2014). «The evolution of seabirds in the Humboldt Current: new clues from the Pliocene of Central Chile». PLOS ONE. 9 (3): e90043. Bibcode:2014PLoSO...990043C. PMC 3951197. PMID 24621560. doi:10.1371/journal.pone.0090043
Ligações externas
- Espécime de Thalassocnus de Lomas del Sauce - Sketchfab
 Media relacionados com Thalassocnus no Wikimedia Commons
Media relacionados com Thalassocnus no Wikimedia Commons O Wikispecies possui informações sobre: Thalassocnus
O Wikispecies possui informações sobre: Thalassocnus