Oryzomys couesi
Oryzomys couesi
| |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Final do Pleistoceno até o presente | |||||||||||||||||||
 | |||||||||||||||||||
| Estado de conservação | |||||||||||||||||||
 Pouco preocupante [1] | |||||||||||||||||||
| Classificação científica | |||||||||||||||||||
| |||||||||||||||||||
| Distribuição geográfica | |||||||||||||||||||
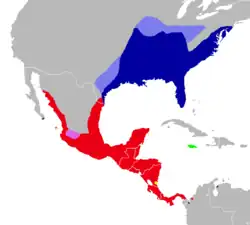 Distribuição de Oryzomys couesi (em vermelho) e outras espécies de Oryzomys.
| |||||||||||||||||||
| Sinónimos | |||||||||||||||||||
Oryzomys couesi, também conhecido popularmente no inglês como "Coues's rice rat", que seria como "rato-do-arroz-de-Coues" no Português, é um roedor semiaquático da família Cricetidae, encontrado desde o sul do Texas, passando pelo México e América Central, até o noroeste da Colômbia. Geralmente habita ambientes úmidos, como pântanos, mas também vive em florestas mais secas e arbustais.
Com um peso de cerca de 43 a 82 g, O. couesi é um rato de tamanho médio a grande. O pelo áspero é amarelado a avermelhado na parte superior e branco a amarelado na parte inferior. As patas traseiras apresentam adaptações para a vida na água, como tufos unguais [en] (como pelos) reduzidos ao redor dos dedos. Possui 56 cromossomos. Há grande variação geográfica em tamanho, proporções, cor e características cranianas.
A espécie é ativa durante a noite e constrói ninhos de vegetação suspensos entre juncos a cerca de 1 m acima do solo. É um excelente nadador e mergulhador, mas também pode escalar vegetação. Como onívoro, alimenta-se de plantas e animais, incluindo sementes e insetos. Reproduz-se ao longo do ano; as fêmeas dão à luz cerca de quatro filhotes após uma gestação de 21 a 28 dias. A espécie pode ser infectada por diversos parasitas e por dois hantavírus.
A espécie foi descrita pela primeira vez em 1877, a primeira de muitas espécies relacionadas da região descritas até a década de 1910. Em 1918, Edward Alphonso Goldman consolidou a maioria em uma única espécie, O. couesi, e em 1960 Raymond Hall [en] uniu este táxon com seu parente dos Estados Unidos, O. palustris, em uma única espécie amplamente distribuída; posteriormente, muitas espécies relacionadas e localizadas mantidas por Goldman também foram incluídas neste táxon.
Após estudos da zona de contato no Texas, onde O. couesi e O. palustris se encontram, que destacaram a distinção entre os dois, eles foram novamente considerados espécies separadas. Desde então, algumas formas periféricas do grupo, como O. antillarum da Jamaica e O. peninsulae [en] da península da Baixa Califórnia, foram reinstauradas como espécies. No entanto, O. couesi, como atualmente constituído, provavelmente é um composto de várias espécies; um estudo de 2010, usando dados de sequência de DNA, encontrou evidências para reconhecer espécies separadas nas áreas do Pacífico e do leste da distribuição de O. couesi e duas espécies adicionais do Panamá e Costa Rica. Geralmente, O. couesi é comum e não apresenta preocupação de conservação, sendo até considerado uma praga em alguns lugares, mas algumas populações estão ameaçadas.
Taxonomia
O. couesi e pelo menos seis espécies mais restritivamente distribuídas formam o grupo O. couesi dentro do gênero Oryzomys. A oitava espécie do gênero, O. palustris, é o único membro de seu próprio grupo[9] (a menos que populações ocidentais sejam classificadas como uma espécie separada, O. texensis).[10] Oryzomys anteriormente incluía muitas outras espécies, que foram reclassificadas em vários estudos culminando em contribuições de Marcelo Weksler e colaboradores em 2006, que removeram mais de quarenta espécies do gênero.[11] Todas são colocadas na tribo Oryzomyini ("ratos-do-arroz"), um grupo diversificado com mais de cem espécies,[12] e em níveis taxonômicos mais altos na subfamília Sigmodontinae da família Cricetidae, junto com centenas de outras espécies de roedores principalmente pequenos.[13]
História
Edward Alston descreveu O. couesi pela primeira vez em 1877, usando três espécimes do México e da Guatemala.[6] Ele nomeou o animal como Hesperomys couesi, colocando-o no gênero agora extinto Hesperomys, e notou semelhanças com o O. palustris (então chamado Hesperomys palustris) e duas espécies agora colocadas em Tylomys.[14] O epíteto específico, couesi, homenageia o naturalista americano Elliott Coues, que realizou muitos estudos sobre roedores norte-americanos.[6] Em 1893, Oldfield Thomas escreveu que a espécie, então colocada no gênero Oryzomys como O. couesi, causou muita confusão sobre sua identidade, pois os três espécimes (um de Cobán, Guatemala, e dois do México) usados por Alston pertenciam, na verdade, a duas ou três espécies diferentes. Ele restringiu o nome couesi ao animal da Guatemala e introduziu o novo nome O. fulgens para um dos animais mexicanos.[7] Várias outras espécies relacionadas foram descritas a partir do início da década de 1890[3] e em 1901 Clinton Hart Merriam uniu muitas delas em um grupo de espécies palustris-mexicanus, que também incluía O. palustris.[15]
Goldman revisou os Oryzomys norte-americanos em 1918 e consolidou muitas formas em uma única espécie (O. couesi), com dez subespécies distribuídas do sul do Texas e oeste do México até a Costa Rica. Ele colocou-a no grupo de O. palustris e várias espécies com distribuições mais limitadas, que ele considerava relacionadas a O. couesi, mas distintas o suficiente para serem classificadas como espécies separadas.[16] Na década de 1930, algumas formas adicionais relacionadas a O. couesi foram descritas.[3] Como então reconhecido, as distribuições do O. palustris, uma espécie dos Estados Unidos, e O. couesi se encontram no sul do Texas. Em 1960, Hall revisou espécimes desta zona de contato e não encontrou motivos para separar as duas espécies; assim, ele reduziu O. couesi a uma subespécie do O. palustris.[17] Outros pesquisadores continuaram essa fusão e, em 1971, todas as outras espécies que Goldman havia colocado no grupo O. palustris foram classificadas como subespécie do O. palustris, junto com O. azuerensis do Panamá, descrito como uma espécie em 1937.[3]
![Distribuição de Oryzomys couesi e espécies relacionadas segundo Goldman (1918): 1, O. couesi couesi; 2, O. c. richmondi; 3, O. c. zygomaticus; 4, O. c. mexicanus; 5, O. c. aztecus; 6, O. c. crinitus; 7, O. c. regillus; 8, O. c. albiventer; 9, O. c. peragrus; 10, O. c. aquaticus; 11, O. gatunensis; 12, O. cozumelae; 13, O. antillarum; 14, O. peninsulae; 15, O. nelsoni.[1]](https://en.wikipedia.org/wiki/File:Oryzomys_distribution_Goldman.png)
Estudos adicionais da zona de contato palustris–couesi no Texas, publicados em 1979, usando mais espécimes e caracteres, indicaram que as duas espécies são, de fato, facilmente distinguíveis ali; portanto, O. couesi passou a ser novamente considerado uma espécie distinta de O. palustris.[3] Posteriormente, algumas das outras formas sinonimizadas sob O. couesi ou O. palustris foram reinstauradas como espécies separadas — O. nelsoni das Ilhas Marías, oeste do México, e O. antillarum da Jamaica.[19] Em 2009, Carleton e Arroyo-Cabrales revisaram os Oryzomys do oeste do México, reafirmaram a distinção de O. nelsoni e reinstauraram O. peninsulae da ponta da península da Baixa Califórnia e O. albiventer do interior do México como espécies.[20] Ainda assim, O. couesi incluía 22 sinônimos,[21] e os pesquisadores escreveram que estudos adicionais sobre O. couesi e espécies relacionadas certamente resultarão no reconhecimento de mais espécies.[22]
Um estudo de 2010 usou dados de sequência de DNA do gene mitocondrial citocromo b (Cytb) e dois marcadores nucleares, o exon 1 do gene da proteína de ligação ao retinóide inter-fotorreceptor (Rbp3) e o intron 2 do gene 1 da álcool desidrogenase (Adh1-I2) para estudar as relações entre populações de O. palustris e O. couesi.[23] Os dados de Cytb colocaram todos os espécimes de O. couesi estudados em um clado irmão de O. palustris; a distância genética média entre os dois grupos foi de 11,30%,[24] muito maior que a distância entre espécies irmãs nos gêneros relacionados Melanomys e Nectomys (7,48% e 7,52%, respectivamente).[25] Dentro do clado O. couesi, duas populações do Panamá e da Costa Rica foram sucessivamente basais aos outros espécimes, que se dividiram em dois grandes subgrupos — um contendo animais do litoral do Pacífico, do oeste do México até El Salvador, e outro contendo ratos do litoral leste, do Texas até a Nicarágua.
As populações do Panamá e da Costa Rica diferiam de 6,53% a 11,93% das outras, e os subgrupos ocidental e oriental diferiam, em média, por 4,41%.[24] Dados dos dois marcadores nucleares de evolução mais lenta, Rbp3 e Adh1-I2, também colocaram exemplos de Oryzomys em dois clados principais, mas não recuperaram os grupos ocidental e oriental de O. couesi como clados separados. Além disso, Adh1-I2 colocou a população da Costa Rica dentro do clado de O. palustris e colocou alguns espécimes ocidentais de O. couesi mais próximos de O. palustris do que do próprio grupo O. couesi.[26] O conjunto de dados combinado apoiou os clados ocidental e oriental dentro de O. couesi e colocou a população da Costa Rica marginalmente mais próxima de O. palustris do que de O. couesi.[27] Usando o conceito de espécie genética, os autores sugeriram que os quatro grupos encontrados dentro de O. couesi deveriam ser reconhecidos como espécies distintas. Se essa sugestão for seguida, o subgrupo oriental manteria o nome O. couesi, o grupo ocidental seria nomeado O. mexicanus, e os nomes apropriados para as espécies do Panamá e da Costa Rica permanecem incertos.[28]
Oeste do México até El Salvador
|
Populações de Oryzomys couesi de Jalisco, oeste do México, até El Salvador formam um único clado Cytb, que Hanson e colegas propuseram reconhecer como a espécie O. mexicanus.[34] Esses animais diferem por 4,4% de O. couesi no sentido estrito,[35] que ocorre ao norte e leste, são separados por cadeias montanhosas deste último, abrigam diferentes espécies de hantavírus, e, segundo Merriam (1901), têm crânios mais robustos, com molares maiores, arcos zigomáticos mais fortes e cristas mais desenvolvidas ao longo das margens da região interorbital do crânio (entre os olhos).[36] Dentro do clado "O. mexicanus", as diferenças de sequência Cytb têm uma média de 2,06%, e os grupos ocidental (Jalisco a Oaxaca) e oriental (Chiapas e El Salvador) formam subgrupos distintos; Hanson e colegas reconheceram esses como subespécies diferentes, mexicanus no oeste e zygomaticus no leste.[37]
Conforme definido por Carleton e Arroyo-Cabrales em 2009, a subespécie O. c. mexicanus ocorre ao longo da costa do Pacífico, de Sonora central até o sudeste de Oaxaca e no interior ao longo de rios até o centro de Michoacán, sul de Morelos, sul de Puebla e noroeste de Oaxaca. Geralmente vive abaixo de 1.000 m de altitude, mas foi encontrado a 1.525 m em Jalisco.[38] Este padrão de distribuição é semelhante ao de outros roedores do oeste do México, como Sigmodon mascotensis, Hodomys alleni, Peromyscus perfulvus e Osgoodomys banderanus, e foi reconhecido como uma zona biogeográfica distinta em algumas revisões.[39] O. c. mexicanus ocorre próximo a três outras espécies do gênero — O. albiventer, O. peninsulae e O. nelsoni — que são maiores e diferentes em algumas proporções e detalhes de coloração.[40]
Joel Asaph Allen descreveu O. mexicanus pela primeira vez como uma espécie completa em 1897 a partir de espécimes de Jalisco. Na mesma publicação, ele também descreveu O. bulleri de Naiarite próximo, mas não os comparou entre si. Merriam adicionou uma segunda espécie de Naiarite, O. rufus, em 1901, observando que era menor e mais avermelhada que mexicanus. Goldman sinonimizou os três como O. c. mexicanus em 1918 e, em 2009, Carleton e Arroyo-Cabrales concordaram, argumentando que as diferenças entre rufus e mexicanus eram relacionadas à idade e dentro da faixa normal de variação do animal.[41]
Outra subespécie, O. c. lambi, foi descrita por Burt em 1934 do litoral central de Sonora,[30] o que estendeu a distribuição da espécie em 640 km na época. Esta forma é cinza-escura, muito mais escura que mexicanus, e tem uma cauda mais curta e jugais mais fracos.[42] Carleton e Arroyo-Cabrales escreveram que é semelhante a mexicanus, mas que pesquisas adicionais são necessárias para determinar se deve ser reconhecida como uma subespécie. Grandes O. couesi do norte de Sinaloa também podem pertencer a esta forma.[39] Goldman escreveu que mexicanus era muito semelhante à subespécie nominal couesi, mas geralmente com pelo mais claro; as partes superiores são mais amareladas que em couesi e as partes inferiores são geralmente brancas, mas podem ser amareladas, a cor normal em couesi.[43]
O. zygomaticus foi descrito pela primeira vez por Merriam em 1901 como uma espécie separada[33] semelhante a mexicanus, mas com os arcos zigomáticos amplamente espalhados e curvados para baixo.[44] Goldman, que o reduziu a uma subespécie de couesi, registrou-o do sudoeste da Guatemala e Chiapas próximo e o descreveu como ligeiramente mais claro que O. c. couesi, mas mais escuro que O. c. mexicanus.[45] Três espécimes do centro de El Salvador têm sequências Cytb semelhantes às de zygomaticus,[46] mas em "The Mammals of El Salvador" (1961), Burt e Stirton registraram apenas a subespécie couesi do país, observando que espécimes de algumas localidades eram ligeiramente mais claros que outros.[47]
Interior do México
Goldman agrupou quatro subespécies de couesi dos planaltos do interior do centro do México — albiventer, crinitus, aztecus e regillus.[51] Três delas (albiventer de Jalisco,[52] crinitus do Distrito Federal,[49] e aztecus de Morelos)[48] foram descritas por Merriam em 1901, e o próprio Goldman descreveu regillus de Michoacán em 1915.[50] Segundo Goldman, aztecus é claro e de dentes grandes,[53] crinitus é grande, escuro e de dentes grandes,[54] regillus é grande e escuro,[51] e albiventer é grande e relativamente claro.[55]
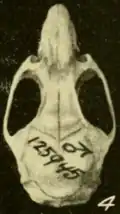
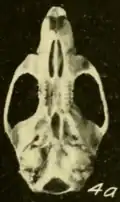
Em sua revisão de 2009 dos Oryzomys do oeste do México, Carleton e Arroyo-Cabrales classificaram O. albiventer como uma espécie separada de mexicanus das terras baixas com base em uma clara diferenciação morfométrica[57] e ofereceram alguns comentários sobre o status de crinitus, regillus e aztecus, incluindo os holótipos das três formas em suas análises morfométricas. Os holótipos de regillus e aztecus estavam na extremidade superior da faixa de variação em sua grande série de mexicanus das terras baixas ocidentais, e crinitus agrupou-se com espécimes de O. peninsulae da ponta da península da Baixa Califórnia. Eles sugeriram que regillus e aztecus podem representar apenas populações robustas das terras altas de mexicanus, mas não puderam excluir a possibilidade de que representem uma espécie diferente. Além disso, sugeriram que crinitus, que ocorre a mais de 2.000 m de altitude no Vale do México, fosse a mesma espécie que peninsulae das terras baixas da península da Baixa Califórnia, eles não aceitaram e recomendaram pesquisas adicionais para determinar as relações de crinitus.[58] Um espécime do interior de Michoacán tem dados Cytb característicos de mexicanus,[59] mas Hanson e colegas não tinham dados para outros Oryzomys do interior do México.[60]
O holótipo da espécie O. fulgens, que Thomas descreveu em 1893, não tem uma localidade mais precisa que "México", mas o Vale do México foi sugerido como sua origem.[61] É uma espécie grande, de pelo áspero, vermelho brilhante,[7] de cauda longa, com um crânio largo com arcos zigomáticos amplamente espalhados.[62] Goldman escreveu que era semelhante a crinitus, mas mais intensamente colorido, e diferia na forma da região interorbital; ele manteve-a como uma espécie separada até investigações adicionais.[63] Carleton e Arroyo-Cabrales notaram que pesquisas arquivísticas podem ainda revelar a origem precisa de O. fulgens, o que poderia estabelecê-la como um nome mais antigo para um dos outros Oryzomys do centro do México.[64]
Texas até Nicarágua
|
Populações de Oryzomys do Texas até a Nicarágua formam um único clado Cytb, com uma divergência média de sequência de 1,28%,[76] e Hanson e colegas propuseram que o nome O. couesi seja restrito a este clado.[28] Essas populações correspondem a duas subespécies reconhecidas por Goldman (O. c. aquaticus e O. c. couesi) e uma forma insular que ele manteve como espécie (O. cozumelae). Outras duas subespécies reconhecidas por Goldman, O. c. richmondi e O. c. peragrus, e uma terceira, O. c. pinicola, descrita após o artigo de Goldman, ocorrem na mesma região, mas não foram estudadas geneticamente.[77]
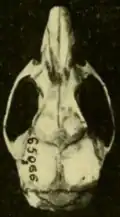
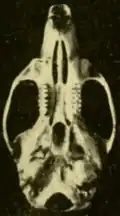
As populações mais ao norte de O. couesi, aquelas no sul do Texas e no Tamaulipas próximo, no México, são classificadas como a subespécie aquaticus, descrita como uma espécie separada, O. aquaticus, em 1891.[78] Aqui, a distribuição de O. couesi encontra a do O. palustris;[79] em partes dos condados de Kenedy, Willacy e Cameron, no Texas, e no extremo nordeste de Tamaulipas, as duas são simpátricas (ocorrem nos mesmos lugares). Na zona de contato, couesi ocorre mais no interior, enquanto o palustris vive ao longo da costa.[80] Em condições experimentais, as duas não se cruzam[81] e a análise genética não mostra evidências de fluxo gênico ou hibridização na natureza.[82] Comparado às populações mais ao sul, aquaticus é maior e mais claro e tem um crânio mais robusto.[83] Espécimes de Tamaulipas são ligeiramente mais escuros que os do Texas.[84] As sequências Cytb de espécimes de aquaticus formam um grupo separado, mas agrupam-se entre espécimes de O. c. couesi de mais ao sul.[85]
A forma peragrus é conhecida de mais ao sul no México, na bacia do rio Verde de San Luis Potosí, o estado de Hidalgo e extremo norte de Veracruz.[86] Fósseis do final do Pleistoceno desta forma foram encontrados na Cueva de Abre, Tamaulipas.[87] Segundo Goldman, é intermediária em cor entre O. c. aquaticus e O. c. couesi, mas tem um crânio semelhante ao de aquaticus.[88]
Goldman uniu populações que vão do norte de Veracruz pelo leste do México, Guatemala, Honduras e Nicarágua até o extremo noroeste da Costa Rica na subespécie nominal, Oryzomys couesi couesi. Ele colocou seis outros nomes como sinônimos completos desta forma, que tem sua localidade-tipo na Guatemala — O. jalapae Allen e Chapman, 1897, de Veracruz; O. j. rufinus Merriam, 1901, de Veracruz; O. teapensis Merriam, 1901, de Tabasco; O. goldmani Merriam, 1901, de Veracruz; O. j. apatelius Eliot, 1904, de Veracruz; e O. richardsoni Allen, 1910, de Nicarágua.[89] Segundo Goldman, a variação individual dentro da subespécie é grande, o que levou ao grande número de sinônimos publicados, mas populações de todas as partes de sua distribuição são essencialmente semelhantes.[90]
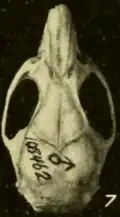
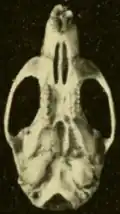
A subespécie O. c. pinicola foi descrita em 1932 de uma crista de pinheiros no oeste de Honduras Britânica (agora Belize); é menor e mais escura que a couesi nominal, que também ocorre em Belize, e tem um crânio mais delicado.[91] Em 1901, Merriam descreveu o Oryzomys da ilha de Cozumel como uma espécie separada, O. cozumelae, e Goldman a manteve como tal por causa de seu grande tamanho, pelo escuro e cauda longa.[92] Em 1965, no entanto, Knox Jones e Timothy Lawlor julgaram as diferenças entre cozumelae e couesi do continente triviais e encontraram que cozumelae estava dentro da faixa de variação das populações continentais de Oryzomys; consequentemente, eles rebaixaram a forma insular a uma subespécie.[93] Mark Engstrom e colegas, escrevendo em 1989, reafirmaram essa conclusão.[94] Para uma forma insular, esta população é altamente variável geneticamente.[95] Em seus dados de sequência Cytb, ela se agrupa entre populações de couesi nominal.[60] O. couesi também é encontrado no "Turneffe Atoll"na costa de Belize[96] e em Roatán na costa de Honduras.[97]
O Oryzomys das terras baixas orientais da Nicarágua foi descrito como uma espécie separada, O. richmondi, por Merriam em 1901, e Goldman o manteve como uma subespécie de O. couesi com base em seu pelo distintamente escuro.[98] Ao revisar os Oryzomys da Nicarágua em 1986, Jones e Engstrom não mantiveram richmondi como separado, porque consideraram a diferença de cor pequena demais para o reconhecimento de subespécies.[99] O. dimidiatus, um Oryzomys pequeno e escuro com partes inferiores cinzentas, ocorre com O. couesi no sudeste da Nicarágua.[100] Segundo Jones e Engstrom, os ratos-arroz da ilha de Ometepe no Lago Nicarágua são distintos por seu crânio grande e medidas externas pequenas, com uma cauda especialmente curta, pelo macio que é laranja-acastanhado acima e amarelado abaixo, e ausência de vacuidades esfenopalatinas (aberturas no teto da fossa mesopterigoide, a lacuna atrás do fim do palato ósseo). Eles consideraram que essa população provavelmente representava uma subespécie separada, mas se recusaram a propor um novo nome porque tinham apenas um espécime adulto.[101] Na Nicarágua, O. couesi ocorre até uma altitude de 1.250 m.[102]
Costa Rica, Panamá e Colômbia
Oryzomys da Costa Rica foram historicamente referidos como O. c. couesi,[105] mas Hanson e colegas descobriram que dois espécimes do Refugio Nacional de Vida Silvestre Mixto Maquenque, nordeste da Costa Rica, diferiam tanto de outros O. couesi (11,93% de divergência de sequência Cytb) quanto O. couesi diferia do O. palustris (11,30%). Eles sugeriram que esses animais representavam uma espécie distinta de O. couesi, mas não conseguiram resolver o nome correto para a espécie porque não puderam examinar amostras de dimidiatus ou richmondi.[106]
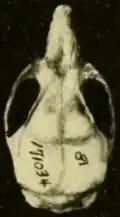
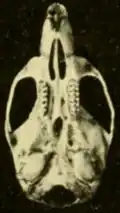
Oryzomys é raro no Panamá.[107] Oryzomys panamenhos foram descritos pela primeira vez por Goldman em 1912, que introduziu o nome Oryzomys gatunensis para um espécime de Gatún [en] na Zona do Canal.[104] Em 1918, Goldman manteve o animal como uma espécie separada, observando que era semelhante a richmondi, mas distinta pelas cristas bem desenvolvidas ao longo das margens da região interorbital, o osso interparietal curto (parte do teto do crânio) e os ossos nasais longos.[108]
Em 1937, Bole descreveu outra espécie de Oryzomys panamenho, O. azuerensis de Paracoté, Veráguas.[103] É uma forma marrom, sem os tons avermelhados das populações próximas, e tem um crânio largo com um rostro (parte frontal) curto e cristas na região interorbital como as de gatunensis.[109] Embora Goldman tenha recomendado a ele que gatunensis e azuerensis fossem tratados como subespécies de couesi, Bole descreveu azuerensis como uma espécie porque não parecia intermediária entre as formas geograficamente mais próximas, gatunensis e couesi, e era separada por uma grande lacuna das populações conhecidas mais próximas de O. couesi no noroeste da Costa Rica e sudeste da Nicarágua.[110] Em uma revisão de 1966 dos mamíferos panamenhos, Charles Handley reduziu tanto gatunensis quanto azuerensis a subespécies de O. palustris (no qual O. couesi estava incluído na época),[111] e quando O. couesi foi reinstaurado como uma espécie separada, essas formas o acompanharam.[3] Espécimes próximos à localidade-tipo de azuerensis diferem por cerca de 7% em suas sequências Cytb de outros O. couesi, o que sugere que podem representar uma espécie separada. No entanto, Hanson e colegas não reinstauraram azuerensis como uma espécie, porque não puderam examinar amostras de gatunensis.[36]
O. couesi foi relatado pela primeira vez na Colômbia em 1987, quando Philip Hershkovitz informou sobre sua ocorrência em Montería em Córdova, noroeste da Colômbia.[112] O espécime colombiano é de cor ocrácea em todo o corpo e, segundo Hershkovitz, quase idêntico aos espécimes da Guatemala, mas distinto pelo lábio superior branco. Ele sugeriu que O. couesi também pode ser descoberto nas terras baixas do Pacífico do Chocó no oeste da Colômbia.[113]
Nomes comuns
Vários nomes comuns foram propostos para O. couesi e os sinônimos atualmente associados a ele. Eliot em 1905[114] e Goldman em 1918 deram nomes comuns separados para cada uma das espécies e subespécies que reconheceram.[115] Muitos autores usaram "rato-arroz-de-Coues" ou alguma variação disso para O. couesi,[116] mas "Oryzomys de Coues" também foi usado.[3]
Descrição
| População | n[fn 3] | Comprimento total | Cauda | Pata traseira |
|---|---|---|---|---|
| Oeste do México até El Salvador | ||||
| San José de Guaymas, Sonora (lambi)[30][fn 4] | 4 | 227 | 113 | 29 |
| Escuinapa, Sinaloa (mexicanus)[43] | 10 | 251.4 (239–273) | 137.4 (127–165) | 28.9 (27–35) |
| Jalisco (mexicanus)[117] | 58 | 245.7 (195–301) | 132.3 (102–160) | 30.2 (26–34) |
| Nayarit (mexicanus)[117] | 62 | 244.8 (210–288) | 125.1 (105–150) | 30.5 (27–33) |
| Nenton, Guatemala (zygomaticus)[45] | 1 | 290 | 152 | 33 |
| El Salvador[118] | 87 | 190–304 | 109–194 | 25–33 |
| Interior do México | ||||
| Morelos (aztecus)[53] | 3 | 307 (297–318) | 161 (154–170) | 34 (33–35) |
| Cidade do México (crinitus)[54] | 2 | 307, 280 | 161, 148 | 37, 35 |
| Michoacán (regillus)[51] | 3 | 308 (285–320) | 168 (155–180) | 35 (34–36) |
| Desconhecido (fulgens)[119] | 1 | 311 | 151 | 37.5[fn 5] |
| Texas até Nicarágua | ||||
| Brownsville, Texas (aquaticus)[83] | 5 | 297 (283–310) | 161 (138–180) | 34.5 (32–38) |
| Rio Verde, San Luis Potosí (peragrus)[88] | 3 | 281 (265–294) | 157 (143–167) | 34 (33–35) |
| Orizaba, Veracruz (couesi)[120] | 7 | 263 (248–294) | 148 (139–174) | 33.1 (32–34.5) |
| Tumbala, Chiapas (couesi)[121] | 4 | 252 (242–265) | 130 (127–135) | 30.7 (30–31) |
| El Cayo, Belize (pinicola)[122] | 18 | 108 (96–128)[fn 6] | 122 (107–146) | 27 (24.6–29) |
| Cozumel (cozumelae)[92] | 6 | 306 (285–327) | 172 (163–177) | 34.3 (33–35.5) |
| Yaruca, Honduras (couesi)[121] | 10 | 267.5 (255–280) | 138 (130–145) | 29.1 (28–32) |
| San Antonio, Nicarágua (couesi)[123] | 25 | 264.9 (242–292) | 135.6 (127–150) | 28.8 (27–31) |
| Sudeste da Nicarágua (richmondi)[98] | 10 | 275.8 (255–295) | 137 (124–151) | 30.9 (29–33.5) |
| Isla de Omotepe, Nicarágua[123] | 1 | 260 | 121 | 30 |
| Costa Rica, Panamá e Colômbia | ||||
| Azuero, Panamá (azuerensis)[109] | 1 | 203 | 107.5 | 30 |
| Gatún, Panamá (gatunensis)[124] | 224 | 115 | 31.5 | |
| Medidas em milímetros e na forma "média (mínimo–máximo)", exceto onde há apenas um ou dois espécimes medidos. | ||||
O. couesi é um rato de tamanho médio a grande[125] com pelo áspero que é amarelado a avermelhado na parte superior, tornando-se mais claro nas laterais e bochechas e mais escuro no traseiro e rosto. As partes inferiores são brancas a amareladas. O pelo é mais curto, mais brilhante e de cor mais intensa do que no O. palustris. O focinho termina de forma abrupta e os olhos moderadamente grandes apresentam um brilho ocular avermelhado. As orelhas pequenas são pretas por fora e o interior é coberto por pelos curtos, cinza a amarelados ou vermelhos. A cauda longa é marrom-escura acima e branca a marrom-clara abaixo.[126] Os pés são longos e robustos.[127] Nas patas dianteiras, os tufos unguais (tufos de pelo nos dedos) estão presentes.[128] Muitas das almofadas nas patas traseiras são reduzidas, assim como os tufos unguais,[129] e pequenas membranas interdigitais podem estar presentes em pelo menos alguns espécimes.[9] Algumas dessas características são adaptações comuns à vida na água em orizomíneos.[130] Como na maioria dos outros orizomíneos, as fêmeas têm oito mamas.[131] O comprimento da cabeça e corpo é de 98 a 142 mm, o comprimento da cauda é de 107 a 152 mm, o comprimento da pata traseira é de 27 a 33 mm, o comprimento da orelha é de 13 a 18 mm, e a massa corporal é de 43 a 82 g.[127] Estudos no Texas e em El Salvador descobriram que os machos são ligeiramente maiores que as fêmeas.[132]
O estômago tem o padrão característico dos sigmodontíneos (unilocular-hemiglandular): não é dividido em duas câmaras por uma incisura angular e a parte frontal (antro) é coberta por um epitélio glandular.[133] A vesícula biliar está ausente, uma sinapomorfia (caráter derivado compartilhado) dos Oryzomyini.[134] O cariótipo inclui 56 cromossomos e um número fundamental de 56 braços autossômicos (2n = 56, FNa = 56). Os autossomos incluem 26 pares de cromossomos acrocêntricos, com um braço longo e um muito curto, e um par submetacêntrico de tamanho médio, com um braço mais curto que o outro.[135] O cromossomo X é acrocêntrico, com um braço longo e um curto, ou subtelocêntrico, com um braço longo e um vestigial.[25] A forma dos cromossomos sexuais foi usada para distinguir o O. palustris de O. couesi, mas não há diferenças consistentes entre os dois.[136]
Como é característico dos Sigmodontinae, O. couesi tem um pênis complexo, com o báculo (osso do pênis) terminando em três dígitos cartilaginosos na ponta.[137] A superfície externa do pênis é majoritariamente coberta por pequenos espinhos penianos [en], mas há uma faixa larga de tecido não espinhoso.[138] A papila (projeção semelhante a um mamilo) na face dorsal (superior) do pênis é coberta por pequenos espinhos, uma característica que O. couesi compartilha apenas com Oligoryzomys e o próximo O. palustris entre os orizomíneos examinados.[139] No processo uretral, localizado no cratera no final do pênis,[140] está presente um processo carnoso (o lóbulo subapical); ele está ausente em todos os outros orizomíneos com pênis estudados, exceto para O. palustris e Holochilus brasiliensis.[141]
Crânio
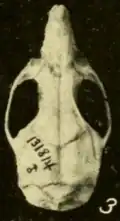
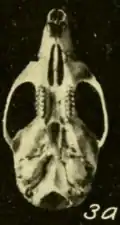
Os ossos nasais e pré-maxilares não se estendem para trás além do ponto onde os ossos lacrimais, frontais e maxilares se encontram.[142] A placa zigomática, a parte frontal achatada do arco zigomático, é larga e desenvolve uma incisura em sua extremidade frontal.[143] A margem posterior da placa está localizada antes do primeiro molar superior.[144] O osso jugal, parte do arco zigomático, é reduzido, como usual em orizomíneos.[145] O forame esfenopalatino, uma abertura na lateral do crânio acima dos molares, é pequeno; é muito maior em O. palustris.[146] A parte mais estreita da região interorbital está voltada para a frente e as bordas são revestidas por prateleiras proeminentes.[147] Os ossos parietais se estendem até as laterais do crânio.[148] O osso interparietal é estreito e em forma de cunha, de modo que os ossos parietal e esquamosal se encontram extensivamente.[149]
Os forames incisivos, aberturas na parte frontal do palato, alcançam para trás entre os molares.[144] O palato é longo, estendendo-se substancialmente além do terceiro molar, a condição usual em orizomíneos.[150] A parte posterior, perto do terceiro molar, é geralmente perfurada por fossas palatais posterolaterais proeminentes, que são recuadas em fossas (depressões).[151] As vacuidades esfenopalatinas estão geralmente ausentes, mas foram relatadas em algumas populações.[152] Não há estrutura alisfenóide, uma extensão do osso alisfenóide que em alguns orizomíneos separa dois forames no crânio.[153] A condição das artérias na cabeça é altamente derivada.[154] O fenestra esquamosal, uma abertura na parte posterior do crânio determinada pela forma do osso esquamosal, está presente.[155] O esquamosal não possui um processo suspensório que contata o tegmen tympani, o teto da cavidade timpânica, um caráter definidor dos orizomíneos.[156] Há algumas aberturas no osso mastóide.[157]
Na mandíbula, o forame mentual, uma abertura logo antes do primeiro molar, abre-se lateralmente, não para cima como em alguns outros orizomíneos.[158] As cristas masseterianas superior e inferior, que ancoram alguns dos músculos mastigatórios, unem-se em um ponto abaixo do primeiro molar e não se estendem para frente além desse ponto.[159] O processo capsular, uma elevação do osso da parte posterior da mandíbula que abriga a extremidade posterior do incisivo, é grande.[160]
Dentes
A fórmula dentária é (um incisivo superior e inferior e três molares superiores e inferiores em cada lado das mandíbulas),[125] como usual em roedores muroides.[161] Os incisivos superiores são opistodontes, com a borda mastigatória localizada atrás do plano vertical dos dentes.[162] Os molares são bunodontes, com as cúspides mais altas que as cristas de conexão, e braquidontes, de coroa baixa, como na maioria dos outros orizomíneos.[163] Muitas cristas acessórias, incluindo o mesolofo nos molares superiores e o mesolofídeo nos molares inferiores, estão presentes, uma característica que O. couesi compartilha com a maioria, mas não todos os outros orizomíneos.[164] Os flexos e flexídeos (vales entre as cúspides e cristas) na face labial (externa) dos molares são fechados por cíngulos (cristas).[165]

Nos primeiro e segundo molares superiores, os flexos não se estendem até a linha média dos molares.[165] O anterocone, a cúspide frontal do primeiro molar superior, não é dividido em dois por uma indentação em sua frente (flexo anteromediano). Uma crista, o anterolofo, está presente atrás da cúspide labial.[166] Como na maioria dos orizomíneos, todos os molares superiores têm uma raiz na face interna (lingual) e duas na face externa (labial); além disso, o primeiro molar superior geralmente tem outra raiz labial pequena.[162]
No primeiro molar inferior, os cônulos labial e lingual do anteroconídeo, a cúspide mais frontal, são separados por uma fosseta anteromediana.[167] O segundo molar inferior possui uma crista, o anterolofídeo, antes das duas cúspides, o protoconídeo e o metaconídeo, que formam a borda frontal do molar em alguns outros orizomíneos.[168] Há uma crista distinta (cíngulo anterolabial) na borda frontal externa (anterolabial) do molar, antes do protoconídeo.[167] O terceiro molar inferior também possui um anterolofídeo e um cíngulo anterolabial.[169] O primeiro molar inferior tem grandes raízes na frente e atrás do dente e duas menores no meio, nas faces labial e lingual. O segundo e terceiro molares inferiores têm duas grandes raízes, uma na frente e uma atrás.[162]
Esqueleto pós-craniano
Como é comum em orizominos, há doze costelas. A primeira costela articula-se com a última cervical (pescoço) e a primeira torácica (tórax), uma sinapomorfia dos Sigmodontinae.[170] Anapófises, processos na parte posterior de uma vértebra, estão ausentes na quinta lombar.[171] Entre a segunda e a terceira vértebras caudais [en], estão presentes arcos hemais (pequenos ossos) com uma borda posterior espinhosa.[172] O forame entepicondilar está ausente, como em todos os membros dos Sigmodontinae; se presente, como em alguns outros roedores, esse forame perfura a extremidade distal (longe) do úmero (osso do braço superior).[173]
Ecologia e comportamento

A distribuição de O. couesi se estende do sul do Texas e centro de Sonora, mas não do planalto central do México, através da América Central até o noroeste da Colômbia;[3] veja em "Taxonomia" para detalhes. A espécie também foi encontrada em depósitos de cavernas do final do Pleistoceno no México e em Honduras.[174] É comum em habitats aquáticos, como pântanos e pequenos riachos, mas também ocorre em florestas e arbustais com cobertura suficiente.[175] Além disso, é encontrada em campos de cana-de-açúcar e arroz.[107] No Texas, ocorre em vegetação de pântano ao longo de "resacas (lagoas em ferradura).[176] Em Veracruz, foi encontrada até mesmo na planície costeira seca entre arbustos.[177] Ocorre desde 2.300 m de altitude até o nível do mar.[125] Em Cozumel, a proporção de juvenis e fêmeas é maior perto de estradas que funcionam como bordas de habitat.[178] Ratos-do-arroz de Cozumel raramente cruzam estradas, o que pode isolar subpopulações na ilha.[179]
O. couesi vive no solo e é semiaquático, passando muito tempo na água,[107] como Alston reconheceu em sua descrição original,[180] mas também é um bom escalador.[181] Um estudo na Costa Rica descobriu que O. couesi é um excelente nadador, mergulhando bem e usando sua cauda para se impulsionar. Provavelmente é capaz de forragear debaixo d'água, o que pode ajudar a diferenciar seu nicho do rato ecologicamente semelhante Sigmodon hirsutus [en], que também nada bem, mas não mergulha.[182] Quando perturbado, O. couesi entra na água e nada para longe.[107] É primariamente ativo durante a noite.[177] A espécie constrói ninhos globulares de vegetação trançada suspensos entre juncos, a cerca de 1 m acima da água ou do solo;[183] no Texas, indivíduos maiores fazem ninhos maiores.[184] Geralmente não faz seus próprios caminhos na vegetação, mas pode usar os de outros roedores.[185]
A densidade populacional varia de 5 a 30 por ha.[1] Em Cozumel, a densidade é de cerca de 14,5 a 16,5 indivíduos por ha, mas mostra grande variação sazonal.[186] No oeste do México, um estudo encontrou uma densidade de 3 indivíduos por ha em floresta de nuvens e 1 por ha em uma área perturbada.[187] Em 24 horas, machos de O. couesi no Texas se deslocam até 153 m e fêmeas até 126 m.[188] A dieta inclui material vegetal, como sementes e partes verdes, e animais, incluindo peixes pequenos, crustáceos, caracóis, insetos como formigas e besouros, e outros invertebrados.[189] Provavelmente se reproduz durante todo o ano e, após uma gestação de 21 a 28 dias,[125] a fêmea produz ninhadas de dois a sete filhotes, com uma média de 3,8.[107] Em 28 fêmeas grávidas da Nicarágua, o tamanho da ninhada variou de um a oito filhotes, com média de 4,4.[123] Os jovens tornam-se reprodutivamente ativos com sete semanas de idade e o ciclo de vida é curto.[125]
A serpente introduzida Boa constrictor preda O. couesi em Cozumel.[179] Parasitas registrados em O. couesi em Veracruz incluem carrapatos, ácaros, pulgas e larvas de moscas não identificados.[185] A pulga Polygenis odiosus foi encontrada em um indivíduo de O. couesi de Cozumel.[190] De dez indivíduos de O. couesi em San Luis Potosí, cinco estavam infectados por vermes nematoides Hassalstrongylus musculi e H. bocqueti, com cerca de 25 vermes por rato, e dois estavam infectados por um ou dois cestoides do gênero Raillietina [en].[191] Os ácaros Eubrachylaelaps circularis e Gigantolaelaps boneti foram encontrados em espécimes de O. couesi em Oaxaca,[192] o piolho sugador Hoplopleura oryzomydis [en] na Nicarágua,[193] os ácaros Laelaps oryzomydis, Echinonyssus microchelae, Ornithonyssus bacoti, Prolistrophorus frontalis e P. bakeri em Colima,[194] e o apicomplexano Eimeria couesii [en] no México.[195] A espécie é infectada por dois hantavírus — vírus Catacamas [en] em Honduras e vírus Playa de Oro no oeste do México — relacionados ao vírus Bayou [en] que infecta o O. palustris, uma causa comum de infecções por hantavírus nos Estados Unidos. No entanto, não há registros de infecções por hantavírus em humanos ligadas aos hantavírus de O. couesi.[196] Em Chiapas a espécie sobrevive facilmente à infecção experimental com vários arbovírus, incluindo o vírus da encefalite equina venezuelana, sugerindo que a espécie pode servir como um reservatório para esse vírus.[197]
Estado de conservação
A IUCN classifica Oryzomys couesi como "Pouco preocupante", devido à sua ampla distribuição, abundância e tolerância a diversos habitats, além de estar presente em várias áreas protegidas. A destruição de habitat, como a drenagem de pântanos, pode ameaçar algumas populações. Em muitas áreas, a espécie é tão comum que é considerada uma praga. Populações persistem até mesmo no Vale do México, conforme evidenciado por uma fotografia publicada em 2006.[198] No entanto, a espécie é listada como ameaçada no Texas, onde sua distribuição é muito limitada, devido à perda de habitat.[199] Em 1979, foi estimado o tamanho da população do rato-do-arroz-de-Coues do Texas em cerca de 15.000 indivíduos. Um estudo de 2001 previu que as mudanças climáticas poderiam levar a população do Texas à extinção, pois não haveria habitats adequados disponíveis.[200] A população de Cozumel diminuiu substancialmente desde meados da década de 1980, possivelmente devido à perturbação do habitat e predação por espécies introduzidas.[186]
Notas
- ↑ A data de publicação do nome de Alston é listada alternativamente na literatura como 1876[2] ou 1877.[3] O nome foi proposto na p. 756 do volume de 1876 dos Proceedings of the Zoological Society of London, mas as partes deste periódico nem sempre foram publicadas no ano do volume.[4] Uma revisão de 2005 confirma que a p. 756 do volume de 1876 foi publicada em 1877.[5]
- ↑ Merriam descreveu Oryzomys cozumelae como novo em ambos os seus artigos de 1901,[68] mas seu artigo nos Proceedings of the Biological Society of Washington foi publicado antes (19 de julho) do que o dos Proceedings of the Washington Academy of Sciences (26 de julho); assim, o primeiro é a publicação original deste nome.[69]
- ↑ Número de espécimes medidos.
- ↑ Apenas médias disponíveis.
- ↑ Com garras.
- ↑ Comprimento da cabeça e corpo em vez de comprimento total.
Referências
- ↑ a b Linzey, A. V.; Timm, R.; Woodman, N.; Matson, J.; Samudio, R. (2017) [errata version of 2016 assessment]. «Oryzomys couesi». Lista Vermelha de Espécies Ameaçadas. 2016: e.T15592A115128044. Consultado em 24 de dezembro de 2019
- ↑ Hanson et al., 2010, p. 336
- ↑ a b c d e f g h Musser & Carleton, 2005, p. 1147
- ↑ Dickinson, 2005, p. 427
- ↑ Dickinson, 2005, tabela 1
- ↑ a b c d Alston, 1877, p. 756
- ↑ a b c d Thomas, 1893, p. 403
- ↑ Hall, 1960, p. 173
- ↑ a b Carleton & Arroyo-Cabrales, 2009, p. 117
- ↑ Hanson et al., 2010, p. 342
- ↑ Weksler et al., 2006, tabela 1
- ↑ Weksler, 2006, p. 3
- ↑ Musser & Carleton, 2005
- ↑ Alston, 1877, pp. 756–757; Musser & Carleton, 2005, p. 1189
- ↑ Merriam, 1901b, p. 275
- ↑ Goldman, 1918, pp. 16, 28–29
- ↑ Hall, 1960, pp. 172–173
- ↑ Goldman, 1918, fig. 3
- ↑ Musser & Carleton, 2005, p. 1152; Weksler et al., 2006
- ↑ Carleton & Arroyo-Cabrales, 2009, p. 94
- ↑ Carleton & Arroyo-Cabrales, 2009, p. 116
- ↑ Carleton & Arroyo-Cabrales, 2009, p. 107
- ↑ Hanson et al., 2010, p. 337
- ↑ a b Hanson et al., 2010, figs.1–2
- ↑ a b Hanson et al., 2010, p. 341
- ↑ Hanson et al., 2010, figs. 1, 3–4
- ↑ Hanson et al., 2010, fig. 5
- ↑ a b Hanson et al., 2010, pp. 342–343
- ↑ Allen, 1897, p. 53
- ↑ a b c Burt, 1934, p. 107
- ↑ Allen, 1897, p. 52
- ↑ Merriam, 1901b, p. 287
- ↑ a b c Merriam, 1901b, p. 285
- ↑ Hanson et al., 2010, fig. 2, p. 343, apêndice I
- ↑ Hanson et al., 2010, tabela 1
- ↑ a b Hanson et al., 2010, p. 343
- ↑ Hanson et al., 2010, fig. 2, tabela 1, apêndice I
- ↑ Carleton & Arroyo-Cabrales, 2009, p. 119
- ↑ a b Carleton & Arroyo-Cabrales, 2009, p. 120
- ↑ Carleton & Arroyo-Cabrales, 2009, pp. 118, 121–122
- ↑ Carleton & Arroyo-Cabrales, 2009, pp. 118–120; Goldman, 1918, p. 34
- ↑ Burt, 1934, p. 108
- ↑ a b Goldman, 1918, p. 34
- ↑ a b Merriam, 1901b, p. 286
- ↑ a b Goldman, 1918, p. 33
- ↑ Hanson et al., 2010, figs. 1–2, apêndice I
- ↑ Burt & Stirton, 1961, p. 61
- ↑ a b Merriam, 1901b, p. 282
- ↑ a b Merriam, 1901b, p. 281
- ↑ a b Goldman, 1915, p. 129
- ↑ a b c Goldman, 1918, p. 37
- ↑ Merriam, 1901b, p. 279
- ↑ a b Goldman, 1918, p. 35
- ↑ a b Goldman, 1918, p. 36
- ↑ Goldman, 1918, p. 38
- ↑ a b c d e Goldman, 1918, plate I
- ↑ Carleton & Arroyo-Cabrales, 2009, p. 118
- ↑ Carleton & Arroyo-Cabrales, 2009, p. 113
- ↑ Hanson et al., 2010, fig. 1, apêndice I
- ↑ a b Hanson et al., 2010, fig. 1
- ↑ Carleton & Arroyo-Cabrales, 2009, pp. 113–114
- ↑ Thomas, 1893, p. 404
- ↑ Goldman, 1918, p. 282
- ↑ Carleton & Arroyo-Cabrales, 2009, p. 114
- ↑ Eliot, 1904, p. 266
- ↑ Allen, 1891, p. 289
- ↑ Merriam, 1901a, p. 103
- ↑ Merriam, 1901a, p. 103; Merriam, 1901b, p. 280
- ↑ Poole & Schantz, 1942, p. 306
- ↑ Merriam, 1901b, p. 288
- ↑ Allen & Chapman, 1897, p. 206
- ↑ Merriam, 1901b, p. 283
- ↑ Murie, 1932, p. 1
- ↑ Allen, 1910, p. 99
- ↑ Merriam, 1901b, p. 284
- ↑ Hanson et al., 2010, fig. 1, tabela 1
- ↑ Goldman, 1918, fig. 3; Murie, 1932, p. 1; Handon et al., 2010, fig. 1
- ↑ Goldman, 1918, pp. 39–40; Hanson et al., 2010, apêndice A
- ↑ Schmidt & Engstrom, 1994, p. 914
- ↑ Schmidt & Engstrom, 1994, pp. 916–917
- ↑ Schmidt & Engstrom, 1994, pp. 915–916
- ↑ Schmidt & Engstrom, 1994, p. 920
- ↑ a b Goldman, 1918, p. 40
- ↑ Baker, 1951, p. 215
- ↑ Hanson et al., 2010, fig. 2
- ↑ Goldman, 1918, p. 39; Jones et al., 1983, p. 379; Hall & Dalquest, 1963, p. 289
- ↑ Dalquest & Roth, 1970, pp. 220, 226
- ↑ a b Goldman, 1918, p. 39
- ↑ Goldman, 1918, pp. 29–30
- ↑ Goldman, 1918, p. 31
- ↑ Murie, 1932, pp. 1–2; Murie, 1935, p. 26
- ↑ a b Goldman, 1918, p. 43
- ↑ Jones & Lawlor, 1965, p. 413
- ↑ Schmidt & Engstrom, 1989, p. 414
- ↑ Vega et al., 2004, p. 210
- ↑ Platt et al., 2000, p. 167
- ↑ Koopman, 1959, p. 237
- ↑ a b Goldman, 1918, p. 32
- ↑ Jones & Engstrom, 1986, p. 10
- ↑ Genoways & Jones, 1971, p. 834
- ↑ Jones & Engstrom, 1986, pp. 10, 12
- ↑ Jones & Engstrom, 1986, p. 7
- ↑ a b Bole, 1937, p. 165
- ↑ a b Goldman, 1912, p. 7
- ↑ Goldman, 1918, p. 31; Harris, 1943, p. 12
- ↑ Hanson et al., 2010, p. 343, apêndice A
- ↑ a b c d e Reid, 2009, p. 207
- ↑ Goldman, 1918, pp. 42–43
- ↑ a b Bole, 1937, p. 166
- ↑ Bole, 1937, p. 167
- ↑ Handley, 1966, p. 781
- ↑ Hershkovitz, 1987, p. 152
- ↑ Hershkovitz, 1987, p. 154
- ↑ Eliot, 1905, pp. 182–186
- ↑ Goldman, 1918, pp. 29–43
- ↑ Eliot, 1905, p. 186; Goldman, 1918, p. 29; Linzey et al., 2008; Duff & Lawson, 2004, p. 54
- ↑ a b Carleton & Arroyo-Cabrales, 2009, tabela 2
- ↑ Burt & Stirton, 1961, p. 60
- ↑ Goldman, 1918, p. 41
- ↑ Goldman, 1918, pp. 30–31
- ↑ a b Goldman, 1918, p. 30
- ↑ Murie, 1932, tabela I
- ↑ a b c Jones & Engstrom, 1986, p. 12
- ↑ Goldman, 1918, p. 42
- ↑ a b c d e Medellín & Medellín, 2006, p. 710
- ↑ Goldman, 1918, p. 29; Reid, 2009, p. 206
- ↑ a b Reid, 2009, p. 206
- ↑ Weksler, 2006, pp. 19, 23, tabela 5
- ↑ Carleton & Musser, 1989, p. 24; Weksler, 2006, pp. 23–25
- ↑ Weksler, 2006, pp. 79, 81
- ↑ Weksler, 2006, p. 17, tabela 5
- ↑ Benson & Gehlbach, 1979, p. 226; Burt & Stirton, 1961, p. 60
- ↑ Weksler, 2006, p. 59
- ↑ Weksler, 2006, pp. 58–59
- ↑ Haiduk et al., 1979, p. 612
- ↑ Schmidt & Engstrom, 1994, p. 916; Hanson et al., 2010, pp. 342–343
- ↑ Weksler, 2006, pp. 55–56
- ↑ Weksler, 2006, pp. 56–57
- ↑ Hooper & Musser, 1964, p. 13; Weksler, 2006, p. 57
- ↑ Hooper & Musser, 1964, p. 7
- ↑ Weksler, 2006, p. 57
- ↑ Weksler, 2006, p. 27, tabela 5
- ↑ Weksler, 2006, pp. 31–32
- ↑ a b Weksler, 2006, p. 32
- ↑ Weksler, 2006, p. 32, tabela 5
- ↑ Schmidt & Engstrom, 1994, p. 917
- ↑ Weksler, 2006, p. 28, tabela 5
- ↑ Weksler, 2006, p. 30
- ↑ Weksler, 2006, p. 31
- ↑ Weksler, 2006, pp. 34–35
- ↑ Weksler, 2006, p. 35
- ↑ Weksler, 2006, p. 37; Carleton & Arroyo-Cabrales, 2009, p. 117; Sánchez et al., 2001, p. 210
- ↑ Weksler, 2006, p. 38, tabela 5
- ↑ Weksler, 2006, p. 37
- ↑ Weksler, 2006, pp. 38–39
- ↑ Weksler, 2006, p. 40
- ↑ Weksler, 2006, pp. 40–41
- ↑ Weksler, 2006, p. 41, tabela 5
- ↑ Weksler, 2006, p. 42
- ↑ Weksler, 2006, pp. 41–42
- ↑ Carleton & Musser, 1984, p. 292
- ↑ a b c Weksler, 2006, p. 43
- ↑ Weksler, 2006, pp. 43–44
- ↑ Weksler, 2006, pp. 44–49
- ↑ a b Weksler, 2006, p. 44
- ↑ Weksler, 2006, pp. 44–45
- ↑ a b Weksler, 2006, p. 49
- ↑ Weksler, 2006, p. 52
- ↑ Weksler, 2006, p. 53
- ↑ Weksler, 2006, p. 52, tabela 5
- ↑ Weksler, 2006, pp. 52–53
- ↑ Weksler, 2006, p. 53; fig. 28
- ↑ Weksler, 2006, p. 54
- ↑ Woodman, 1995, tabela 1, p. 225; Woodman & Croft, 2005, tabela 4; Dalquest & Roth, 1970, pp. 220, 226
- ↑ Reid, 2009, p. 207; Hall & Dalquest, 1963, p. 287
- ↑ Schmidly & Davis, 2004, p. 381; Benson & Gehlbach, 1979, p. 228
- ↑ a b Hall & Dalquest, 1963, p. 287
- ↑ Fuentes-Montemayor et al., 2009, p. 865
- ↑ a b Vega et al., 2004, p. 217
- ↑ Alston, 1877, p. 757
- ↑ Murie, 1935, p. 26; Reid, 2009, p. 207
- ↑ Cook et al., 2001
- ↑ Benson & Gehlbach, 1979, p. 227; Reid, 2009, p. 207; Schmidly & Davis, 2004, p. 281
- ↑ Benson & Gehlbach, 1979, p. 227
- ↑ a b Hall & Dalquest, 1963, p. 288
- ↑ a b Vega et al., 2004, p. 218
- ↑ Vázquez et al., 2000, tabela 1
- ↑ Benson & Gehlbach, 1979, p. 226
- ↑ Medellín & Medellín, 2006, p. 710; Reid, 2006, p. 303; 2009, p. 207
- ↑ Eckerlin, 2005, p. 155
- ↑ Underwood et al., 1986
- ↑ Estébanes-González & Cervantes, 2005, p. 27
- ↑ Emerson, 1971, p. 334
- ↑ Estébanes-González et al., 2011, tabela 1
- ↑ Barnard et al., 1971, p. 1294
- ↑ Chu et al., 2008; Milazzo et al., 2006
- ↑ Deardoff et al., 2009, p. 519; 2010, p. 350
- ↑ Medellín & Medellín, 2006, p. 710; Carleton & Arroyo-Cabrales, 2009, p. 113
- ↑ Schmidly & Davis, 2004, p. 381
- ↑ Cameron & Scheel, 2001, p. 676
Literatura citada
- Allen, J.A. 1891. Notes on new or little-known North American mammals, based on recent additions to the collection of mammals in the American Museum of Natural History. Bulletin of the American Museum of Natural History 3:263–310.
- Allen, J.A. 1897. Further notes on mammals collected in Mexico by Dr. Audley C. Buller, with descriptions of new species. Bulletin of the American Museum of Natural History 9:47–57.
- Allen, J.A. 1910. Additional mammals from Nicaragua. Bulletin of the American Museum of Natural History 28:87–115.
- Allen, J.A. and Chapman, F.M. 1897. On a collection of mammals from Jalapa and Las Vigas, state of Vera Cruz, Mexico. Bulletin of the American Museum of Natural History 9:197–208.
- Alston, E.R. 1877 ["1876"]. On two new species of Hesperomys. Proceedings of the Zoological Society of London 1876(4):755–757.
- Alston, E.R. 1882. Biologia centrali-americana. Mammalia. R.H. Porter, 220 pp.
- Baker, R.H. 1951. Mammals from Tamaulipas, Mexico. University of Kansas Publications, Museum of Natural History 5:207–215.
- Barnard, W.P., Ernst, J.V. and Stevens, R.O. 1971. Eimeria palustris sp. n. and Isospora hammondi sp. n. (Coccidia: Eimeriidae) from the marsh rice rat, Oryzomys palustris (Harlan). The Journal of Parasitology 57(6):1293–1296.
- Benson, D.E. and Gehlbach, F.R. 1979. Ecological and taxonomic notes on the rice rat (Oryzomys couesi) in Texas. Journal of Mammalogy 60(1):225–228.
- Bole, B.J. Jr. 1937. Annotated list of the mammals of the Mariato River District of the Azuero Peninsula. Scientific Publications of the Cleveland Museum of Natural History 7:140–196.
- Burt, W.H. 1934. A new rice rat (Oryzomys) from Sonora, Mexico. Proceedings of the Biological Society of Washington 47:107–108.
- Burt, W.H. and Stirton, R.A. 1961. The mammals of El Salvador. Miscellaneous Publications of the Museum of Zoology, University of Michigan 117:1–69.
- Cameron, G.N. and Scheel, D. Getting warmer: Effect of global climate change on distribution of rodents in Texas. Journal of Mammalogy 82(3):652–680.
- Carleton, M.D. and Arroyo-Cabrales, J. 2009. Review of the Oryzomys couesi complex (Rodentia: Cricetidae: Sigmodontinae) in Western Mexico. Bulletin of the American Museum of Natural History 331:94–127.
- Carleton, M.D. and Musser, G.G. 1984. Muroid rodents. pp. 289–379 in Anderson. S. and Jones, J.K. Jr. (eds.). Orders and families of Recent mammals of the world. John Wiley and Sons, New York, 686 pp.
- Chu, Y.-K., Owen, R.D., Sánchez-Hernández, C., Romero-Almarez, M. de L. and Jonsson, C.B. 2008. Genetic characterization and phylogeny of a hantavirus from Western Mexico. Virus Research 131:180–188.
- Cook, W.M., Timm, R.M. and Hyman, D.E. 2001. Swimming ability in three Costa Rican dry forest rodents. Revista de Biología Tropical 49(3–4):1177–1181.
- Dalquest, W.W. and Roth, E. 1970. Late Pleistocene mammals from a cave in Tamaulipas, Mexico. The Southwestern Naturalist 15(2):217–230.
- Deardorff, E.R., Forrester, N.L., Travassos da Rosa, A.P., Estrada-Franco, J.G., Navarro-Lopez, R., Tesh, R.B. and Weaver, S.C. 2009. Experimental infection of potential reservoir hosts with Venezuelan equine encephalitis virus, Mexico. Emerging Infectious Diseases 15(4):519–525.
- Deardorff, E.R., Forrester, N.L., Travassos da Rosa, A.P., Estrada-Franco, J.G., Navarro-Lopez, R., Tesh, R.B. and Weaver, S.C. 2010. Short report: Experimental infections of Oryzomys couesi with sympatric arboviruses from Mexico. American Journal of Tropical Medicine and Hygiene 82(2):350–353.
- Dickinson, E.C. 2005. The Proceedings of the Zoological Society of London, 1859–1900: an exploration of breaks between calendar years of publication. Journal of Zoology 266(4):427–430.
- Duff, A. and Lawson, A. 2004. Mammals of the World: A checklist. New Haven: A & C Black, 312 pp. ISBN 0-7136-6021-X
- Eckerlin, R.P. 2005. Fleas (Siphonaptera) of the Yucatan Peninsula (Campeche, Quintana Roo, and Yucatan), Mexico. Caribbean Journal of Science 41(1):152–157.
- Eliot, D.G. 1904. Descriptions of apparently new species and subspecies of mammals and a new generic name proposed. Field Columbian Museum Zoological Series 3:263–270.
- Eliot, D.G. 1905. A checklist of mammals of the North American continent, the West Indies and the neighboring seas. Field Columbian Museum Zoological Series 6:1–761.
- Emerson, K.C. 1971. Records of a collection of Mallophaga and Anoplura from Nicaraguan mammals. Journal of the Kansas Entomological Society 44(3):332–334.
- Engstrom, M.D., Schmidt, C.A., Morales, J.C. and Dowler, R.C. 1989. Records of mammals from Isla Cozumel, Quintana Roo, Mexico. The Southwestern Naturalist 34(3):413–415
- Estébanes-González, M.L. and Cervantes, F.A. 2005. Mites and ticks associated with some small mammals in Mexico. International Journal of Acarology 31(1):23–37. (inscrição necessária)
- Estébanes-González, M.L., Sánchez-Hernández, C., Romero-Almaraz, M. de L. and Schnell, G.D. 2011. Ácaros parásitos de roedores de Playa de Oro, Colima, México Arquivado em 2016-03-03 no Wayback Machine. Acta Zoológica Mexicana 27(1):169–176 (em espanhol).
- Fuentes-Montemayor, E., Cuarón, A.D., Vázquez-Domínguez, E., Benítez-Malvido, J., Valenzuela-Galván, D. and Andresen, E. 2009. Living on the edge: roads and edge effects on small mammal populations. Journal of Animal Ecology 78:857–865.
- Genoways, H.H. and Jones, J.K. Jr. 1971. Second specimen of Oryzomys dimidiatus. Journal of Mammalogy 52:833–834.
- Goldman, E.A. 1912. Descriptions of twelve new species and subspecies of mammals from Panama. Smithsonian Miscellaneous Collections 56(36):1–11.
- Goldman, E.A. 1915. Five new rice rats of the genus Oryzomys from Middle America. Proceedings of the Biological Society of Washington 27:127–130.
- Goldman, E.A. 1918. The rice rats of North America. North American Fauna 43:1–100.
- Haiduk, M.W., Bickham, J.W. and Schmidly, D.J. 1979. Karyotypes of six species of Oryzomys from Mexico and Central America. Journal of Mammalogy 60(3):610–615.
- Hall, E.R. 1960. Oryzomys couesi only subspecifically different from the marsh rice rat, Oryzomys palustris. The Southwestern Naturalist 5(3):171–173.
- Hall, E.R. and Dalquest, W.W. 1963. The mammals of Veracruz. University of Kansas Publications, Museum of Natural History 14:165–362.
- Handley, C.O. Jr. 1966. Checklist of the mammals of Panama. pp. 753–795 in Wenzel, R.L. and Tipton, V.J. (eds.). Ectoparasites of Panama. Chicago: Field Museum of Natural History, xii + 861 pp.
- Hanson, J.D., Indorf, J.L., Swier, V.J. and Bradley, R.D. 2010. Molecular divergence within the Oryzomys palustris complex: evidence for multiple species (abstract only). Journal of Mammalogy 91(2):336–347.
- Harris, W.P. Jr. 1943. A list of mammals from Costa Rica. Occasional Papers of the Museum of Zoology, University of Michigan 476:1–15.
- Hershkovitz, P. 1987. First South American record of Coues' marsh rice rat, Oryzomys couesi. Journal of Mammalogy 68(1):152–154.
- Hooper, E.T. and Musser, G.G. 1964. The glans penis in Neotropical cricetines (Family Muridae) with comments on classification of muroid rodents. Miscellaneous Publications of the University of Michigan Museum of Zoology 123:1–57.
- Jones, J.K. Jr. and Engstrom, M.D. 1986. Synopsis of the rice rats (genus Oryzomys) of Nicaragua. Occasional Papers, The Museum, Texas Tech University 103:1–23.
- Jones, J.K. Jr. and Lawlor, T.E. 1965. Mammals from Isla Cozumel, México, with description of a new species of harvest mouse. University of Kansas Publications, Museum of Natural History 16:409–419.
- Jones, J.K. Jr., Carter, D.C. and Webster, W.D. 1983. Records of mammals from Hidalgo, Mexico. The Southwestern Naturalist 28(3):378–380.
- Kays, R.W. and Wilson, D.E. 2000. Mammals of North America. Princeton and Oxford: Princeton University Press, 240 pp. ISBN 0-691-07012-1
- Koopman, K.F. 1959. The zoogeographical limits of the West Indies. Journal of Mammalogy 40(2):236–240.
- Linzey, A. V.; Timm, R.; Woodman, N.; Matson, J.; Samudio, R. (2016). «Oryzomys couesi». Lista Vermelha de Espécies Ameaçadas. 2016: e.T15592A115128044. Consultado em 24 de dezembro de 2019
- Medellín, X.L. and Medellín, R.A. 2006. Oryzomys couesi (Alston, 1877). pp. 709–710 in Ceballos, G. and Oliva, G. (eds.). Los mamíferos silvestres de México. Mexico City: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad and Fondo de Cultura Económica, 986 pp. ISBN 978-970-9000-30-6
- Merriam, C.H. 1901a. Six new mammals from Cozumel Island, Yucatan. Proceedings of the Biological Society of Washington 14:99–104.
- Merriam, C.H. 1901b. Synopsis of the rice rats (genus Oryzomys) of the United States and Mexico. Proceedings of the Washington Academy of Sciences 3:273–295.
- Milazzo, M.L., Cajimat, M.N., Hanson, J.D., Bradley, R.D., Quintana, M., Sherman, C., Velásquez, R.T. and Fulhorst, C.F. 2006. Catacamas virus, a hantaviral species naturally associated with Oryzomys couesi (Coues' oryzomys) in Honduras. American Journal of Tropical Medicine and Hygiene 75(5):1003–1010.
- Murie, A. 1932. A new Oryzomys from a pine ridge in British Honduras. Occasional Papers of the Museum of Zoology, University of Michigan 245:1–3.
- Murie, A. 1935. Mammals from Guatemala and British Honduras. Miscellaneous Publications of the Museum of Zoology, University of Michigan 26:1–30.
- Musser, G.G. and Carleton, M.D. 2005. Superfamily Muroidea. pp. 894–1531 in Wilson, D.E. and Reeder, D.M. (eds.). Mammal Species of the World: a taxonomic and geographic reference. 3rd ed. Baltimore: The Johns Hopkins University Press, 2 vols., 2142 pp. ISBN 978-0-8018-8221-0
- Platt, S.G., Rainwater, T.R., Miller, B.W. and Miller, C.M. 2000. Notes on the mammals of Turneffe Atoll, Belize. Caribbean Journal of Science 36(1–2):166–168.
- Poole, A.J. and Schantz, V.S. 1942. Catalog of the type specimens of mammals in the United States National Museum, including the Biological Surveys collection. Bulletin of the United States National Museum 178:1–705.
- Reid, F. 2006. A Field Guide to Mammals of North America, North of Mexico. 4th ed. Houghton Mifflin Harcourt, 579 pp. ISBN 978-0-395-93596-5
- Reid, F. 2009. A Field Guide to the Mammals of Central America and Southeast Mexico. 2nd edition. Oxford University Press US, 346 pp. ISBN 978-0-19-534322-9
- Sánchez H., J., Ochoa G., J. and Voss, R.S. 2001. Rediscovery of Oryzomys gorgasi (Rodentia: Muridae) with notes on taxonomy and natural history. Mammalia 65:205–214.
- Schmidly, D.J. and Davis, W.B. 2004. The mammals of Texas. 2nd edition. University of Texas Press, 501 pp. ISBN 978-0-292-70241-7
- Schmidt, C.A. and Engstrom, M.D. 1994. Genic variation and systematics of rice rats (Oryzomys palustris species group) in southern Texas and northeastern Tamaulipas, Mexico. Journal of Mammalogy 75(4):914–928.
- Thomas, O. 1893. Notes on some Mexican Oryzomys. Annals and Magazine of Natural History (6)11:402–405.
- Underwood, H.T., Owen, J.G. and Engstrom, M.D. 1986. Endohelminths of three species of Oryzomys (Rodentia: Cricetidae) from San Luis Potosi, Mexico. The Southwestern Naturalist 31(3):410–411.
- Vázquez, L.B., Medellín, R.A. and Cameron, G.N. 2000. Population and community ecology of small rodents in montane forest of western Mexico. Journal of Mammalogy 81(1):77–85. (inscrição necessária)
- Vega, R., Vázquez-Domínguez, E., Mejía-Puente, A. and Cuaro, A.D. 2004. Unexpected high levels of genetic variability and the population structure of an island endemic rodent (Oryzomys couesi cozumelae). Biological Conservation 137:210–222.
- Weksler, M. 2006. Phylogenetic relationships of oryzomyine rodents (Muroidea: Sigmodontinae): separate and combined analyses of morphological and molecular data. Bulletin of the American Museum of Natural History 296:1–149.
- Weksler, M., Percequillo, A.R. and Voss, R.S. 2006. Ten new genera of oryzomyine rodents (Cricetidae: Sigmodontinae). American Museum Novitates 3537:1–29.
- Woodman, N. 1995. Morphological variation between Pleistocene and Recent samples of Cryptotis (Insectivora: Soricidae) from the Yucatan Peninsula, Mexico. Journal of Mammalogy 76(1):223–231.
- Woodman, N. and Croft, D. 2005. Fossil shrews from Honduras and their significance for late glacial evolution in body size (Mammalia: Soricidae: Cryptotis). Fieldiana Geology 51:1–30.