Melão (cetáceos)

O melão é uma massa de tecido adiposo encontrada na testa de todas as baleias dentadas.[1][2] Ele foca e modula as vocalizações do animal, atuando como uma lente sonora. É, portanto, um órgão-chave envolvido na comunicação e na ecolocalização.
Descrição
O melão é estruturalmente parte do aparelho nasal e compreende a maior parte do tecido entre o espiráculo e a ponta do focinho. A função do melão não é completamente compreendida, mas os cientistas acreditam que ele seja um componente bioacústico, proporcionando um meio de focar os sons usados na ecolocalização e criando uma similaridade entre as características de seu tecido e a água circundante, de modo que a energia acústica possa fluir da cabeça para o ambiente com a menor perda de energia. Anteriormente, hipotetizava-se que o melão tinha funções em mergulhos profundos e flutuabilidade, mas essas ideias não são mais consideradas plausíveis por cetologistas.[3]
A composição variada do melão cria um gradiente de velocidade sonora que refrata o som de forma direcional. Os sons também refletem no crânio e nos sacos aéreos que cercam o melão.[4]
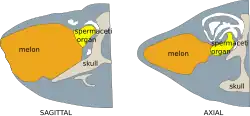
O tamanho do melão não está relacionado à profundidade máxima de mergulho em baleias dentadas. As características particulares do melão provavelmente têm mais a ver com a filogenia dos odontocetos, ou seja, as relações taxonômicas ao longo do tempo evolutivo. Em algumas espécies, os melões são mais especializados do que em outras. A cachalote possui o maior nariz de qualquer animal no mundo. A maior parte desse nariz é composta por duas grandes estruturas gordurosas, o órgão espermaceti [en] e o "junk". O junk é estruturalmente idêntico ao melão (homólogo a ele). O melão não é homólogo ao órgão de espermaceti.[1][5] Pesquisas sobre a expressão de genes como o da proteína MYH16 mostram que ele se origina do músculo masseter.[6]
Composição
O melão é uma mistura de triglicerídeos e ésteres de cera [en]. A composição exata varia ao longo do melão. Tipicamente, o núcleo interno do melão tem um maior conteúdo de cera do que as partes externas e conduz o som mais lentamente. Esse gradiente refrata o som e o foca como uma lente.
Os lipídios no melão não podem ser digeridos pelo animal, pois são metabolicamente tóxicos. Um golfinho faminto mantém um melão robusto mesmo que o resto de seu corpo esteja emaciado.[1] Os lipídios no melão tendem a ter menor peso molecular e serem mais saturados do que a gordura.
Os melões dos Delphinidae [en] (golfinhos oceânicos) e Physeteroidea (cachalotes) possuem uma quantidade significativa de ésteres de cera, enquanto os dos Phocoenidae (toninhas) e Monodontidae (narvais e baleias-beluga) contêm pouco ou nenhuma cera.[7] A velocidade do som no melão é mais baixa nos Delphinidae, Phocoenidae e Monodontidae, intermediária nos Ziphiidae (baleias-de-bico) e mais alta nos Physeteridae e Platanistidae (golfinhos de rio do sul da Ásia).[8]
Baleias-piloto
O melão das baleia-pilotos (Globicephala) é uma mistura de ésteres de cera e triglicerídeos. O núcleo interno do melão contém cerca de 33% de ésteres de cera, enquanto a camada externa contém cerca de 5%. A maioria das gorduras é saturada.[9]
Cachalote-pigmeu
No cachalote-pigmeu (Kogia breviceps), o melão consiste em uma camada externa e um núcleo interno, que geralmente possui uma proporção maior de ésteres de cera do que a camada externa.[10]
Atrás do melão, há um órgão em forma de cornucópia que muitos cientistas chamam de "órgão espermaceti". Este órgão é diferente em forma e composição do órgão de espermaceti da cachalote.
| Melão externo | Melão interno | Órgão de espermaceti | ||
|---|---|---|---|---|
| Conteúdo lipídico (peso) | 15–91% | 74–94% | 92–96% | |
| Composição lipídica |
Ésteres de cera | 8–46% | 40–90% | 84–99% |
| Triglicerídeos | 54–92% | 10–69% | 1–16% | |
| Número médio de carbonos |
Ésteres de cera | 32–35 | 29–32 | 28–29 |
| Triglicerídeos | 47–51 | 41–46 | 45 | |
Cachalote
As cachalotes possuem dois grandes sacos cheios de óleo, empilhados um sobre o outro: o órgão espermaceti dorsal ou caixa de espermaceti e o "junk",[11] nomeado porque os baleeiros o consideravam inútil para a extração de óleo de espermaceti. O junk originou-se do melão dos odontocetos.[12] Ele contém compartimentos de óleos cerosos separados por paredes de tecido conjuntivo.[11] Juntos, o órgão espermaceti e o junk adicionam direcionalidade e amplitude aos cliques do biossonar.[13][14]
Baleia-beluga
O melão da baleia-beluga é único, pois a baleia pode alterar a forma do melão à vontade.[15] Essas mudanças na forma provavelmente alteram o tamanho, a forma, a direção e a composição de frequência do feixe de ecolocalização.
Referências
- ↑ a b c Cranford, Ted W.; Amundin, Mats; Norris, Kenneth S. (junho de 1996). «Functional morphology and homology in the odontocete nasal complex: Implications for sound generation». Journal of Morphology. 228 (3): 223–285. PMID 8622183. doi:10.1002/(SICI)1097-4687(199606)228:3<223::AID-JMOR1>3.0.CO;2-3
- ↑ Harper, C.J.; McLellan, W.A.; Rommel, S.A.; Gay, D.M.; Dillaman, R.M.; Pabst, D.A. (julho de 2008). «Morphology of the melon and its tendinous connections to the facial muscles in bottlenose dolphins (Tursiops truncatus)» (PDF). Journal of Morphology. 269 (7): 820–839. Bibcode:2008JMorp.269..820H. PMID 18473369. doi:10.1002/jmor.10628
- ↑ Cranford, Ted W. (outubro de 1999). «The sperm whale's nose: Sexual selection on a grand scale?». Marine Mammal Science. 15 (4): 1133–1157. doi:10.1111/j.1748-7692.1999.tb00882.x
- ↑ Marine Mammal Biology: An Evolutionary Approach. pg 153
- ↑ Cranford, Ted W.; Mckenna, Megan F.; Soldevilla, Melissa S.; Wiggins, Sean M.; Goldbogen, Jeremy A.; Shadwick, Robert E.; Krysl, Petr; St. Leger, Judy A.; Hildebrand, John A. (abril de 2008). «Anatomic Geometry of Sound Transmission and Reception in Cuvier's Beaked Whale (Ziphius cavirostris)». The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology. 291 (4): 353–378. PMID 18228579. doi:10.1002/ar.20652

- ↑ Takeuchi, Hayate; Matsuishi, Takashi Fritz; Hayakawa, Takashi (2024). «A tradeoff evolution between acoustic fat bodies and skull muscles in toothed whales». Gene. 901. PMID 38224921. doi:10.1016/j.gene.2024.148167
- ↑ Litchfield, Carter; Ackman, R. G.; Sipos, J. C.; Eaton, C. A. (setembro de 1971). «Isovaleroyl triglycerides from the blubber and melon oils of the beluga whale (Delphinapterus leucas)». Lipids. 6 (9): 674–681. PMID 5141491. doi:10.1007/BF02531529
- ↑ Litchfield, Carter; Greenberg, Anne J.; Caldwell, David K.; Caldwell, Melba C.; Sipos, J. C.; Ackman, R. G. (abril de 1975). «Comparative lipid patterns in acoustical and non-acoustical fatty tissues of dolphins, porpoises and toothed whales». Comparative Biochemistry and Physiology Part B: Comparative Biochemistry. 50 (4): 591–597. PMID 1122741. doi:10.1016/0305-0491(75)90095-4
- ↑ Wedmid, Yuri; Litchfield, Carter; Ackman, R. G.; Sipos, J. C.; Eaton, C. A.; Mitchell, E. D. (dezembro de 1973). «Heterogeneity of lipid composition within the cephalic melon tissue of the pilot whale (Globicephala melaena)». Biochimica et Biophysica Acta (BBA) - Lipids and Lipid Metabolism. 326 (3): 439–447. PMID 4776442. doi:10.1016/0005-2760(73)90144-6
- ↑ a b Karol, R.; Litchfield, C.; Caldwell, D. K.; Caldwell, M. C. (1978). «Compositional topography of melon and spermaceti organ lipids in the pygmy sperm whale Kogia breviceps: Implications for echolocation». Marine Biology. 47 (2): 115–123. Bibcode:1978MarBi..47..115K. doi:10.1007/BF00395632
- ↑ a b Clarke, Malcolm R. (fevereiro de 1978). «Structure and Proportions of the Spermaceti Organ in the Sperm Whale». Journal of the Marine Biological Association of the United Kingdom (em inglês). 58 (1): 1–17. Bibcode:1978JMBUK..58....1C. ISSN 1469-7769. doi:10.1017/S0025315400024371
- ↑ Heyning, John E.; Mead, James G. (1990), «Evolution of the Nasal Anatomy of Cetaceans», ISBN 978-1-4899-0860-5, Boston, MA: Springer US, Sensory Abilities of Cetaceans, pp. 67–79, doi:10.1007/978-1-4899-0858-2_5, consultado em 21 de agosto de 2025
- ↑ Møhl, B.; Wahlberg, M.; Madsen, P. T.; Miller, L. A.; Surlykke, A. (1 de janeiro de 2000). «Sperm whale clicks: Directionality and source level revisited». The Journal of the Acoustical Society of America. 107 (1): 638–648. Bibcode:2000ASAJ..107..638M. ISSN 0001-4966. PMID 10641672. doi:10.1121/1.428329
- ↑ Møhl, Bertel (1 de junho de 2001). «Sound transmission in the nose of the sperm whale Physeter catodon. A post mortem study». Journal of Comparative Physiology A (em inglês). 187 (5): 335–340. ISSN 1432-1351. PMID 11529477. doi:10.1007/s003590100205
- ↑ «Beluga Whale wiggling its Melon at Vancouver Aquarium». YouTube. 31 de maio de 2010. Cópia arquivada em 21 de dezembro de 2021