Gordodon
| Gordodon | |
|---|---|

| |
| Espécime holótipo (NMMNH P-70796) | |
| Classificação científica | |
| Reino: | Animalia |
| Filo: | Chordata |
| Clado: | Synapsida |
| Família: | †Edaphosauridae |
| Gênero: | †Gordodon Lucas et al., 2018 |
| Espécies: | †G. kraineri
|
| Nome binomial | |
| †Gordodon kraineri Lucas et al., 2018
| |
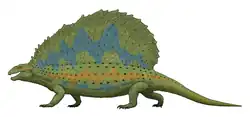
Gordodon[1] (significando "dente gordo", em referência aos seus grandes dentes frontais semelhantes a incisivos) é um gênero extinto de sinapsídeo não mamífero que viveu durante o Permiano Inferior (Cisuraliano) na região que hoje corresponde ao condado de Otero, Novo México. Pertence à família herbívora Edaphosauridae, e inclui apenas uma espécie, a espécie-tipo Gordodon kraineri. Gordodon destaca-se entre os sinapsídeos primitivos por sua dentição, que se assemelha à dos mamíferos modernos, diferente dos dentes simples e uniformes, semelhantes aos de lagartos, de outros sinapsídeos herbívoros primitivos. Possuía grandes dentes frontais semelhantes a incisivos, seguidos por um espaço pronunciado (diastema) e uma fileira curta de dentes em forma de pinos na parte posterior da boca. Gordodon também apresentava um pescoço relativamente longo para um sinapsídeo primitivo, com vértebras alongadas no pescoço e nas costas. Como outros membros de sua família, tinha uma vela dorsal alta formada pelas espinhas neurais ósseas de suas vértebras. Essas espinhas também possuíam protuberâncias ósseas, uma característica comum entre os membros de Edaphosauridae, mas as de Gordodon eram únicas por serem mais delgadas, semelhantes a espinhos, e distribuídas aleatoriamente ao longo das espinhas. Estima-se que fosse relativamente pequeno, com cerca de 1 metro de comprimento (excluindo a cauda) e aproximadamente 34 kg de peso.
Os maxilares e dentes únicos de Gordodon entre os sinapsídeos primitivos sugerem que ele foi um dos primeiros tetrápodes herbívoros a se especializar em uma alimentação seletiva de frutificações vegetais ricas em nutrientes e pobres em fibras, como sementes e estruturas semelhantes a frutos. Foi também um dos primeiros tetrápodes a exibir uma dentição e aparelho alimentar tão especializados, evoluindo apenas alguns milhões de anos após o surgimento dos primeiros herbívoros tetrápodes obrigatórios no registro fóssil. Antes da descoberta de Gordodon, os sinapsídeos não mamíferos herbívoros com dentes igualmente complexos eram os cinodontes, semelhantes a mamíferos, que surgiram 95 milhões de anos depois, durante o Triássico.
Descoberta e nomenclatura

O único fóssil conhecido de Gordodon foi descoberto em março de 2013 por Ethan Schuth, estudante de geologia da Universidade de Oklahoma, durante uma excursão de campo. O espécime foi encontrado exposto ao longo de uma estrada próximo à cidade de Alamogordo, no condado de Otero, Novo México, em estratos geológicos identificados como pertencentes à base da formação Bursum [en]. A equipe contatou o Museu de História Natural e Ciência do Novo México (sigla em inglês: NMMNH) em Albuquerque, que coletou o espécime entre 2013 e 2014, catalogando-o como NMMNH P-70796. Durante a escavação, o crânio foi acidentalmente cortado verticalmente, deixando uma marca de 6,25 mm de largura através dos maxilares e da órbita.[1]
A descrição do espécime foi publicada em 2018 pelos paleontólogos Spencer G. Lucas [en], Larry F. Rinehart e Matthew D. Celeskey, que o nomearam como o holótipo de um novo gênero e espécie, Gordodon kraineri. O nome genérico deriva do espanhol "gordo", que significa "gordo", e do sufixo grego "-odon", significando "dente gordo", em referência aos característicos dentes frontais grandes. O nome também faz alusão a "Alamogordo", a cidade próxima ao local da descoberta. O nome específico kraineri homenageia Karl Krainer, geólogo da Universidade de Innsbruck, reconhecendo seu extenso trabalho em paleontologia e geologia no Novo México.[1]
O único espécime conhecido de Gordodon é um esqueleto incompleto, mas articulado, exposto principalmente no lado direito, que inclui a porção frontal do animal: o crânio e os maxilares inferiores, cinco vértebras cervicais no pescoço, quatro vértebras torácicas completas das costas e partes das altas espinhas neurais de outras 12 vértebras não visíveis na placa, costelas, partes de ambas as escápulas e clavículas nos ombros, e dois dígitos parciais, provavelmente da mão. Embora pequeno, a sutura das vértebras e do ombro sugere que o espécime não era um juvenil.[1]
O espécime-tipo de Gordodon foi descoberto em uma posição estratigráfica baixa na formação Bursum, em um local que Lucas e colegas chamaram de "localidade dos Edaphosauridae". Esse local está a apenas 3 metros acima da base da formação, que foi datada aproximadamente como pertencente ao Cisuraliano norte-americano, geralmente considerado do Permiano Inferior, equivalente ao estágio Asseliano globalmente definido.[2] Gordodon está na fronteira entre o subperíodo Pensilvânico do Carbonífero Superior e o Permiano Inferior, com cerca de 299 milhões de anos, podendo ser considerado do Pensilvânico com base em definições alternativas para a base do Permiano (por exemplo, bioestratigrafia de conodontes).[3] O fóssil foi encontrado em fácies fluviais de arenitos cinza-oliva depositados por canais fluviais.[1]
Descrição
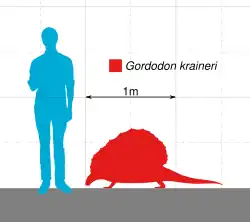
Gordodon era um membro da família Edaphosauridae relativamente pequeno, com um comprimento estimado da cabeça aos quadris (comprimento pré-sacral) de cerca de 1 metro, sem incluir a cauda. Estima-se que pesava apenas 34 kg, menos da metade do tamanho da maioria das espécies de Edaphosaurus. Comparado a membros posteriores de sua família, as costelas de Gordodon eram muito menos curvadas, sugerindo que não tinha o torso em forma de barril como Edaphosaurus, mas sim um torso mais estreito e reto, semelhante ao dos membros da família Sphenacodontidae, como Dimetrodon. As vértebras são relativamente esbeltas, com uma característica única entre os membros da família Edaphosauridae: duas quilhas na parte inferior. As vértebras do pescoço têm centros retangulares e alongados, enquanto as das costas são mais quadradas. Além de parte da cintura escapular, os únicos ossos dos membros conhecidos de Gordodon são dois dígitos incompletos, provavelmente de uma das mãos. Esses dígitos são longos e esguios, com garras curvas e pontiagudas nas extremidades, mais semelhantes aos de Remigiomontanus [en] do que aos dígitos mais robustos de Edaphosaurus. Outros aspectos da anatomia de Gordodon permanecem desconhecidos, já que é conhecido apenas por um único espécime parcial.[1][4]
Crânio
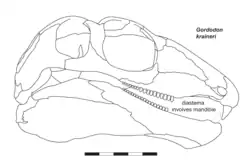
O crânio de Gordodon é relativamente grande para um membro da família Edaphosauridae, medindo 159 mm de comprimento da frente para trás. Embora, como outros indivíduos de sua família, a cabeça ainda seja proporcionalmente pequena em relação ao corpo. O crânio é ligeiramente arqueado para cima ao longo de seu comprimento, com um focinho relativamente longo e estreito, aproximadamente igual em comprimento à porção do crânio atrás dos olhos. As órbitas (cavidades oculares) são grandes e arredondadas, com uma "sobrancelha" óssea proeminente acima delas, formada pelos ossos pré-frontal, frontal e pós-frontal. A única abertura atrás dos olhos (a fenestra temporal, ponto de fixação dos músculos mandibulares), característica dos sinapsídeos, é aproximadamente quadrada e menor que a órbita, embora seja notavelmente mais alta do que larga.[1]
Os maxilares e dentes de Gordodon são uma de suas características mais distintivas. Os únicos dentes na frente do maxilar são um par de grandes dentes semelhantes a incisivos nas extremidades dos maxilares superior e inferior. Apenas os "incisivos" superiores estão preservados, embora um par oposto no maxilar inferior seja inferido por um alvéolo dentário aberto no dente direito visível. Esses dentes parecem triangulares e pontiagudos quando vistos de lado, mas provavelmente eram retangulares e em forma de cinzel quando vistos de frente. São os únicos dentes na pré-maxila (ossos frontais do maxilar superior), e atrás deles há um diastema longo e curvado para cima, um espaço proeminente nos maxilares entre os dentes frontais e traseiros, formado pela maxila (o principal osso do maxilar superior). Após o diastema, há 18 dentes pequenos em forma de pinos em cada maxila, com 8 dentes ligeiramente maiores na frente e 6 menores atrás (não está claro se a transição entre eles era abrupta ou gradual devido a danos no fóssil). Os dentes no maxilar inferior são semelhantes em tamanho e forma aos do maxilar superior, e há um diastema oposto na frente, embora mais suavemente curvado. Como Edaphosaurus, Gordodon tinha densas baterias de dentes pequenos em forma de pinos nas superfícies internas dos maxilares inferiores, dispostas em placas dentárias distintas. Essas provavelmente correspondiam a um conjunto oposto nos ossos pterigoides no céu da boca, embora essa área não seja visível no único espécime conhecido. Dentes palatais muito pequenos (<1 mm de diâmetro) são visíveis no vômer (um osso no palato), formando dois grupos alongados ao longo da linha média do diastema entre os dentes das bochechas e os "incisivos". O maxilar inferior é notavelmente mais profundo na parte traseira, com dentes relativamente rasos e uma sínfise mandibular não fundida (onde os dois maxilares inferiores se conectam na frente), que é apenas ligeiramente inclinada para baixo, formando um leve "queixo".[1]
Vela

Como outros membros da família Edaphosauridae, Gordodon possuía uma grande vela sustentada por espinhas neurais extremamente alongadas em suas vértebras, que se estendiam pelo pescoço e costas. Curiosamente, a vela de Gordodon apresenta uma morfologia de transição entre os membros anteriores da família e Edaphosaurus. Diferentemente dos membros anteriores, as espinhas neurais são mais espessas e comprimidas lateralmente, quase como lâminas em suas extremidades, embora não sejam tão robustas quanto as de Edaphosaurus. Além disso, Gordodon não possui uma espinha alongada em seu áxis (a segunda vértebra do pescoço), ao contrário de Ianthasaurus, de modo que a vela começa apenas sobre a terceira vértebra cervical, como em Edaphosaurus. As espinhas neurais também apresentam numerosas protuberâncias ósseas, como as de Edaphosaurus. No entanto, não formam fileiras organizadas de barras transversais grossas e geralmente simétricas, como visto em Edaphosaurus. Em vez disso, as protuberâncias de Gordodon são finas e pontiagudas, como espinhos, e estão distribuídas aleatoriamente de forma assimétrica em ambos os lados, sem um padrão discernível. Inusitadamente, as espinhas neurais continuam a aumentar substancialmente em comprimento até a 12ª espinha, enquanto em Edaphosaurus e em Ianthasaurus a altura da espinha se estabiliza na 8ª espinha. Da mesma forma, a espinha neural mais alta é a 16ª, de modo que a vela atinge o pico mais para trás do que em Edaphosaurus (12ª espinha). Combinados, esses fatores conferem a Gordodon uma vela com uma inclinação muito mais acentuada em comparação com as velas mais semicirculares de Edaphosaurus e de Ianthasaurus.[1]
Classificação
Gordodon pertence à família Edaphosauridae, um grupo de sinapsídeos majoritariamente onívoros e herbívoros com vela dorsal dentro do clado Eupelycosauria, com o qual compartilha uma cabeça caracteristicamente pequena, tamanho reduzido dos dentes ao longo das margens do maxilar, uma articulação maxilar abaixo da fileira de dentes e espinhas neurais altas com protuberâncias laterais. Gordodon distingue-se de todos os outros membros de Edaphosauridae por sua dentição variada, ou heterodonte, incluindo seus "incisivos" em forma de cinzel, diastema e dentes das bochechas em forma de pinos. Também pode ser distinguido por uma sutura relativamente curta entre os ossos nasais e a maxila, vértebras cervicais e dorsais relativamente graciosas com duas quilhas na parte inferior e protuberâncias em forma de espinhos distribuídas aleatoriamente em suas espinhas neurais.[1]
Uma análise filogenética foi realizada por Lucas e colegas em 2018 para determinar as relações de Gordodon com outros membros de sua família. Outra análise das relações dos membros da família Edaphosauridae foi publicada por Spindler e colegas em 2019, incluindo espécies recém-descritas da família. Ambas as análises posicionaram Gordodon de forma semelhante em relação aos outros membros da família, mais derivado que Ianthasaurus e Glaucosaurus, mas menos que Lupeosaurus [en] e Edaphosaurus.[1][4]
O cladograma de Spindler et al. (2019) é mostrado simplificado abaixo:

| Edaphosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Ambas as análises observaram que Gordodon ajudou a resolver a porção "intermediária" da árvore de Edaphosauridae, que em análises anteriores era uma região instável com relações entre espécies pouco definidas.[5] Gordodon combina características de Edaphosauridae anteriores, como centros vertebrais mais longos e um focinho relativamente longo, com traços mais derivados, como as numerosas protuberâncias ósseas nas espinhas neurais. Da mesma forma, exibe vários traços intermediários entre edafossaurídeos anteriores e derivados, incluindo um crânio geral menor, comprimento de focinho reduzido e uma mandíbula mais profunda em comparação com Edaphosauridae anteriores. A relativa completude de Gordodon permitiu comparações com Edaphosauridae anteriores e derivados que não eram conhecidos por materiais sobrepostos (por exemplo, crânios versus pós-crânio), esclarecendo as relações entre eles.[1][4]
História evolutiva
Diastemas são encontrados nos maxilares superiores de outros sinapsídeos primitivos, incluindo os predadores Kenomagnathus [en] e Tetraceratops.[6][7] No entanto, Gordodon é, até o momento, o único sinapsídeo herbívoro não terapsídeo a possuir um verdadeiro diastema. Também é o único sinapsídeo fora dos cinodontes a ter um diastema dentário oposto ao do maxilar superior. O diastema e os dentes heterodontes de Gordodon, semelhantes aos dos mamíferos, evoluíram independentemente dos terapsídeos mais derivados, incluindo mamíferos, indicando que a evolução dos maxilares e dentes mamíferos não foi um processo linear, com diastematas funcionais evoluindo várias vezes em sinapsídeos primitivos, incluindo os membros de Edaphosauridae. O diastema de Gordodon provavelmente originou-se de um pequeno espaço na fileira de dentes superiores dos membros de Edaphosauridae primitivos (um "diastema inicial"), como Ianthasaurus, formado pela sobreposição das fixações do pré-maxilar e da maxila.[6]
Dentro de Edaphosauridae, Gordodon também indica que pelo menos dois estilos distintos de herbivoria evoluíram cedo na história da família: uma dieta especializada de baixa fibra em Gordodon; e uma alimentação geral de plantas ricas em fibras em membros mais derivados de Edaphosauridae, como Edaphosaurus. No entanto, ainda não está claro se ambos os estilos descendem de herbívoros menos especializados de baixa fibra ou se cada um evoluiu de um ancestral mais onívoro, semelhante a Ianthasaurus.[5] A idade do Permiano Inferior de Gordodon também sugere que os herbívoros de Edaphosauridae se diversificaram cedo em sua evolução, aumentando a possibilidade de uma maior diversidade ecológica dentro de Edaphosauridae ainda a ser descoberta.[1]
Paleobiologia
Como outros membros de Edaphosauridae, Gordodon era herbívoro, embora sua dentição única sugira que se alimentava de alimentos diferentes de seus parentes. Em sua descrição por Lucas e colegas em 2018, eles sugeriram que as características especializadas de seus maxilares e dentes indicavam que Gordodon era um alimentador mais seletivo, uma interpretação corroborada por Spindler em 2020.[6] Essa interpretação baseia-se em características como seu focinho mais estreito, "incisivos" especializados e diastema, que juntos teriam permitido a Gordodon cortar e processar seletivamente a vegetação em pedaços menores antes de engoli-los. Essas características são encontradas em mamíferos vivos conhecidos por serem alimentadores seletivos.[8] Esse método de alimentação poderia ter permitido a Gordodon consumir matéria vegetal mais rica em nutrientes e pobre em fibras do que outros membros de sua família, especialmente frutificações (tecidos vegetais não vegetativos, como sementes e frutos). Embora frutos verdadeiros sejam produzidos apenas por angiospermas, que ainda não haviam evoluído, estruturas semelhantes a frutos são encontradas em gimnospermas, incluindo sementes carnosas e pinhas de coníferas, gnetófitas e Cycadales.[9][10] A caixa torácica relativamente estreita e "de lados retos" de Gordodon pode apoiar essa interpretação, já que uma dieta de baixa fibra requer menos tempo para digestão, e assim Gordodon não precisaria de um intestino grande e arredondado para fermentar vegetação fibrosa.[1]
Lucas e colegas (2018) inferiram que Gordodon usava seus grandes "incisivos" para cortar alimentos, semelhante a alguns roedores e coelhos modernos, colocando os alimentos no diastema antes de serem passados para trás para serem moídos pelas placas dentárias. Os dentes simples e em forma de pinos da maxila e do dente funcionavam para cortar a vegetação. A articulação maxilar baixa e desalinhada permite que os dentes superiores e inferiores se ocluam ao longo de todo o seu comprimento, conferindo a Gordodon uma mordida mais forte e esmagadora, em vez de cortante. Além disso, a articulação maxilar é frouxa e possui uma articulação arredondada e esférica, permitindo um movimento palinal do maxilar inferior (onde o maxilar é deslizado para trás contra o maxilar superior) para moagem e corte adicionais dos alimentos. Os dentes das "bochechas" de Gordodon não são especializados para esse propósito, ao contrário dos mamíferos vivos, e a mastigação provavelmente era realizada pelas placas dentárias. Isso seria uma forma relativamente mais complexa de mastigação em comparação com o movimento palinal mais limitado observado no maxilar inferior de Edaphosaurus.[1][11] No entanto, a função exata do diastema e o movimento dos maxilares em relação às placas dentárias nos maxilares superior e inferior é difícil de determinar, como observado pelo paleontólogo Frederik Spindler em 2020, que sugeriu ainda o papel de uma língua especializada. Portanto, os mecanismos exatos dos maxilares e dentes durante o processamento de alimentos de Gordodon, assim como de outros membros de sua família, permanecem desconhecidos.[6]
Paleoecologia
As fácies sedimentares da formação Bursum indicam que Gordodon habitava um ambiente de planície costeira próximo à costa. Durante a deposição da formação, o habitat alternava entre ambientes terrestres, de praia e marinhos, devido a ciclos de elevação e queda do nível do mar, indicando sua proximidade com a costa. A maioria dos organismos conhecidos da formação Bursum são marinhos, com restos particularmente abundantes de invertebrados, incluindo uma variedade de gastrópodes, ostracodos, equinodermos, braquiópodes e bivalves, além de pequenos briozoários e foraminíferos.[2][12] Restos fragmentários de vertebrados de outras partes da formação, fora da localidade de Edaphosauridae, incluem restos de peixes, membros da ordem Temnospondyli aquáticos, membros da ordem Embolomeri [en], membros do clado Diadectomorpha herbívoros, membros da família Caseidae, membros da família Sphenacodontidae predadores e possivelmente outros membros de Edaphosauridae.[13]
Referências
- ↑ a b c d e f g h i j k l m n o Spencer G., Lucas; Rinehart, Larry; Celeskey, Matthew D. (novembro de 2018). «The oldest specialized tetrapod herbivore: A new eupelycosaur from the Permian of New Mexico, USA». Palaeontologia Electronica. 21.3.39A (3): 1–42. doi:10.26879/899

- ↑ a b Krainer, K.; Vachard, D.; Lucas, S. G. (2003). «Microfacies and microfossil assemblages (smaller foraminifers, algae, pseudoalgae) of the Hueco Group and Laborcita Formation (Upper Pennsylvanian-lower Permian), south-central New Mexico, USA». Rivista Italiana di Paleontologia e Stratigrafia. 109 (1): 3–36. doi:10.13130/2039-4942/5491
- ↑ Davydov, V. I.; Glenister, B. F.; Spinosa, C.; Ritter, S. M.; Chernykh, V. V.; Wardlaw, B. R.; Snyder, W. S. (1998). «Proposal of Aidaralash as Global Stratotype Section and Point (GSSP) for base of the Permian System». Episodes. 21 (1): 11–17. doi:10.18814/epiiugs/1998/v21i1/003
- ↑ a b c Spindler, Frederik; Voigt, Sebastian; Fischer, Jan (2019). «Edaphosauridae (Synapsida, Eupelycosauria) from Europe and their relationship to North American representatives». PalZ. 94: 125–153. doi:10.1007/s12542-019-00453-2
- ↑ a b Mazierski, D. M.; Reisz, R. R. (2010). «Description of a new specimen of Ianthasaurus hardestiorum (Eupelycosauria: Edaphosauridae) and a re-evaluation of edaphosaurid phylogeny». Canadian Journal of Earth Sciences. 47 (6): 901–912. Bibcode:2010CaJES..47..901M. doi:10.1139/e10-017
- ↑ a b c d Spindler, F. (2020). «A faunivorous early sphenacodontian synapsid with a diastema». Palaeontologia Electronica. 23 (1). doi:10.26879/1023
- ↑ Spindler, F. (2019). «The skull of Tetraceratops insignis (Synapsida, Sphenacodontia)». Palaeovertebrata. 43 (1): e1. doi:10.18563/pv.43.1.e1
- ↑ Pérez-Barbería, E. J.; Gordon, I. J. (1999). «The functional relationship between feeding type and jaw and cranial morphology in ungulates». Oecologia. 118 (2): 157–165. Bibcode:1999Oecol.118..157P. PMID 28307690. doi:10.1007/s004420050714
- ↑ Lovisetto, A.; Guzzo, F.; Busatto, N.; Casadoro, G. (2013). «Gymnosperm B-sister genes may be involved in ovule/seed development and, in some species, in the growth of fleshy fruit-like structures». Annals of Botany. 112 (3): 535–544. PMC 3718214. PMID 23761686. doi:10.1093/aob/mct124
- ↑ Hall, J. A.; Walter, G. H. (2014). «Relative Seed and Fruit Toxicity of the Australian Cycads Macrozamia miquelii and Cycas ophiolitica: Further Evidence for a Megafaunal Seed Dispersal Syndrome in Cycads, and Its Possible Antiquity». Journal of Chemical Ecology. 40 (8): 860–868. PMID 25172315. doi:10.1007/s10886-014-0490-5
- ↑ Reisz, Robert R. (2006). «Origin of dental occlusion in tetrapods: Signal for terrestrial vertebrate evolution?». Journal of Experimental Zoology Part B: Molecular and Developmental Evolution. 306B (3): 261–277. PMID 16683226. doi:10.1002/jez.b.21115
- ↑ Krainer, K.; Lucas, S. G. (2013). «The Pennsylvanian–Permian Bursum Formation in central New Mexico». The Carboniferous–Permian Transition in Central New Mexico: New Mexico Museum of Natural History and Science, Bulletin. 59: 143–160
- ↑ Harris, S. K.; Lucas, S. G.; Berman, D. S.; Henrici, A. C. (2004). «Vertebrate fossil assemblage from the Upper Pennsylvanian Red tanks member of the Bursum Formation, Lucero uplift, central New Mexico». Carboniferous-Permian Transition at Carrizo Arroyo, Central New Mexico: New Mexico Museum of Natural History and Science, Bulletin. 25: 267–283
Ligações externas
 Media relacionados com Gordodon no Wikimedia Commons
Media relacionados com Gordodon no Wikimedia Commons
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||


.jpg)