Dendrosenecio
Dendrosenecio
| |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Classificação científica | |||||||||||||||||||||
| |||||||||||||||||||||
| Espécie-tipo | |||||||||||||||||||||
| Dendrosenecio johnstonii | |||||||||||||||||||||
| Distribuição geográfica | |||||||||||||||||||||
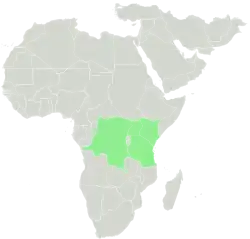 Distribuição natural do género.
| |||||||||||||||||||||
| Espécies | |||||||||||||||||||||
| Sinónimos[1][2] | |||||||||||||||||||||



Dendrosenecio é um género de plantas com flor pertencente à família Asteraceae,[5][6][7] recentemente segregado do género Senecio,[1] no qual formava o subgénero Dendrosenecio.[1] Os seus membros, pequenas árvores com morfologia muito distinta, são nativos das zonas de maior altitude de dez grupos montanhosos da região equatorial da África Oriental,[8] onde constituem um elemento conspícuo da flora.
Descrição
Os membros deste género apresentam um hábito caracterizado pela presença de uma roseta gigante terminal no ápice de um robusto caule lenhoso. Quando florescem, as flores formam uma grande inflorescência terminal. Concomitantemente, normalmente surgem dois a quatro ramos laterais. Como resultado, as plantas mais velhas têm a aparência de candelabros do tamanho de postes telefónicos, cada ramo com uma roseta de folhas terminal.[8]
Espécies
O género Dendrosenecio varia geograficamente entre cadeias montanhosas e altitudinalmente em cada montanha. Tem havido desacordo entre os botânicos quanto às populações de Dendrosenecio que merecem ser reconhecidas como espécies e quais devem ser relegadas para o estatuto de subespécies ou variedades. A lista seguinte apresenta a listagem de maior consenso:[8]
- Dendrosenecio adnivalis (Stapf) E.B.Knox (1993)
- Dendrosenecio battiscombei (R.E.Fr. & T.C.E.Fr.) E.B.Knox (1993)
- Dendrosenecio brassiciformis (R.E.Fr. & T.C.E.Fr.) Mabb. (1986)
- Dendrosenecio cheranganiensis (Cotton & Blakelock) E.B.Knox (1993)
- Dendrosenecio elgonensis (T.C.E.Fr.) E.B.Knox (1993)
- Dendrosenecio erici-rosenii (R.E.Fr. & T.C.E.Fr.) E.B.Knox (1993)
- Dendrosenecio johnstonii (Oliv.) B.Nord. (1978)
- Dendrosenecio keniensis (Baker f.) Mabb. (1986)
- Dendrosenecio keniodendron (R.E.Fr. & T.C.E.Fr.) B.Nord. (1978)
- Dendrosenecio kilimanjari (Mildbr.) E.B.Knox (1993)
- Dendrosenecio meruensis (Cotton & Blakelock) E.B.Knox (1993)
Distribuição
Várias espécies do género Senecio são encontradas em todo o mundo como ervas daninhas comuns à beira da estrada, mas em nenhum outro lugar, exceto nas terras altas de África, exibem formas arbóreas tão grandes.
Os membros do género Dendrosenecio encontram-se na zona alpina das montanhas da África Oriental equatorial – Monte Kilimanjaro e Monte Meru na Tanzânia, Monte Quénia, a Cordilheira de Aberdare e Colinas Cherangani no Quénia, o Monte Elgon na fronteira entre o Uganda e o Quénia, os Montes Rwenzori na fronteira entre o Uganda e a República Democrática do Congo (RDC), os Montes Virunga nas fronteiras do Ruanda, do Uganda e da República Democrática do Congo, e os Montes Mitumba (Monte Kahuzi e Monte Muhi) no leste da República Democrática do Congo.
Com exceção de Dendrosenecio eric-rosenii, que ocorre em várias montanhas do Rift Albertino (Montanhas Rwenzori, Virunga e Mitumba), e de Dendrosenecio battiscombei e Dendrosenecio keniodendron, que são partilhadas pelo Monte Quénia e pela Cordilheira de Aberdare, as espécies estão individualmente confinadas a uma única cordilheira. Em várias das áreas de distribuição, encontram-se espécies diferentes, ou subespécies, em altitudes diferentes.
As áreas de distribuição natural das diferentes espécies são as seguintes:[8]
| Cadeia | Kilimanjaro | Meru | Monte Kenya | Aberdares | Cherangani | Elgon | Ruwenzori | Virunga | Mitumba |
|---|---|---|---|---|---|---|---|---|---|
| Grande altitude | Dendrosenecio kilimanjari ssp. cottonii | Dendrosenecio keniodendron | D. keniodendron | D. elgonensis ssp. barbatipes | |||||
| Altitude intermédia | D. kilimanjari ssp. kilimanjari | D. meruensis | D. keniensis | D. brassiciformis | D. cherangiensis ssp. dalei | D. elgonensis ssp. elgonensis | D. adnivalis (two subspecies) | D. eric-rosenii ssp. alticola | |
| Baixa altitude | D. johnstonii | D. battiscombei | D. battiscombei | D. cherangiensis ssp. cherangiensis | D. eric-rosenii ssp. eric-rosenii | D. eric-rosenii ssp. eric-rosenii | D. eric-rosenii ssp. eric-rosenii |
Evolução e adaptação
As montanhas da África Central e da África Oriental são um modelo quase ideal para estudar a especiação e a adaptação nas plantas. As montanhas elevam-se muito acima das planícies e planaltos circundantes,[3] suficientemente elevadas para alcançar altitudes acima da linha das árvores[10] e formar ilhas do céu.[3] Estes picos predominantemente vulcânicos simplificam ainda mais o modelo devido à sua idade e disposição em torno da bacia do Lago Vitória e à proximidade do equador.[3]
As espécies encontradas no Monte Quénia são, de longe, o melhor modelo de variação altitudinal. A espécie D. keniodendron é a espécie que cresce nas altitudes mais elevadas, a D. keniensis encontra-se nas altitudes mais baixas da área de distribuição onde a espécie cresce e a D. battiscombei cresce nas mesmas altitudes que a D. keniensis mas nos ambientes mais húmidos. As outras montanhas que não são suficientemente altas para ter uma "grande no topo" têm as duas, uma espécie nas terras mais secas e outra nos ambientes mais húmidos, ou apenas uma onde o ambiente não é tão extremo. Esta simplificação funciona muito bem como introdução à distribuição das espécies gigantes de Dendrosenecio da África Oriental, com uma exceção, o Kilimanjaro, que tem uma espécie que ocorre no topo e apenas uma espécie que ocorre em baixas altitudes; as subespécies e variedades ocorrem nos ambientes mais húmidos.
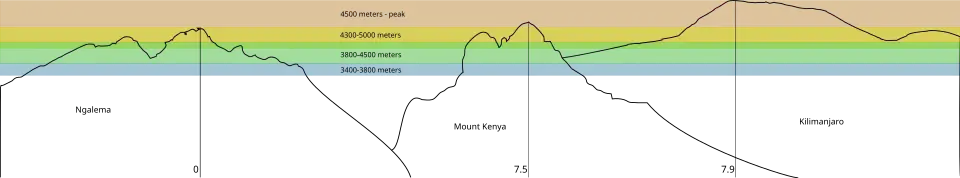
Cada montanha tem um gradiente vertical de precipitação e diferentes flutuações diárias de temperatura.[11] O Monte Kilimanjaro, com 5 895 m de altitude no seu cume, o Monte Quénia com 5 199 m e as Ruwenzori com 5 109 m são as três montanhas mais altas de África, cada uma suficientemente alta para suportar diferents faixas de zonas vegetativas baseadas na altitude.[12] Em consequência, cada montanha oferece o seu próprio conjunto de habitats isolados, dispostos verticalmente.[3]
Localizadas entre de 50 a 1000 km em torno da linha do equador, nessas montanhas as flutuações ambientais ocorrem como eventos diários,[3] resultando em dias quentes e noites frias que são uniformes ao longo do ano[11] ou, como Karl Olov Hedberg descreveu a situação, verão todos os dias, inverno todas as noites.[13] Para além das variáveis ambientais simplificadas, estas regiões de montanhas são facilmente circunscritas, o que simplifica a análise biogeográfica, uma vez que a sua idade e disposição em torno da bacia do Lago Vitória facilitam a separação dos efeitos do tempo e da posição.[3]
Zonas de vegetação
Nas altitudes entre 3 400 e 4 500 m podem ser encontrados alguns dos exemplos mais extremos de adaptação, que incluem:
- Rosetas de folhas maciças em que o desenvolvimento das folhas ocorre numa grande roseta apical;
- Armazenamento de água na medula do caule (paquicaulia);
- Isolamento do caule através da retenção de folhagem murcha e morta;
- Secreção e retenção de fluidos polissacarídeos nucleantes de gelo (um anticongelante natural);
- Movimento foliar nictinástico (as folhas fecham-se quando faz frio).[3]
Em altitudes inferiores a 3 400 m as flutuações diárias de temperatura são menos extremas, a temperatura média diária aumenta constantemente e as formas de crescimento e a ecologia do Dendrosenecio reflectem a maior influência dos factores bióticos (como a competição pela luz) sobre os factores abióticos (como a geada nocturna).[3]
- Altitudes de 3 400 a 3 800 m
O nome da região Afroalpina foi proposto por Lucien Leon Hauman em 1955.[14] Existe um limite nítido a 3 400 metros de altitude (3 000 metros no lado norte) que separa a floresta da zona alpina inferior,[10] o ambiente é uma charneca (vegetação rasteira em solos ácidos) e é aqui que os Dendrosenecio começam a crescer entre as gramíneas de montanha e os juncos.[15] D. keniensis cresce nesta região no Monte Quénia. Uma variedade ou subespécie de D. johnstonii ocorre nesta faixa de altitude nas três montanhas mais altas.
- Altitudes de 3 800 a 4 500 m
As charnecas superiores ocorrem nestas altitudes e é nessas regiões que a maioria das populações de Dendrosenecio brassica se instala nas três montanhas, convivendo com os os matos esclerófilos de arbustos anões resistentes à seca.
- Altitudes de 4 300 a 5 000 m
Florestas de Dendrosenecio, onde cada montanha tem a sua própria variedade. D. keniensis no Monte Quénia, D. kilimanjari no Monte Kilimanjaro e outras espécies em cada uma das cadeias montanhosas.
- Altitudes acima de 4 500 m— (picos)
As populações de Dendrosenecio começam a diminuir. O Monte Quénia tem a menor vegetação nas suas partes superiores devido às suas baixas temperaturas.
A dispersão e estabelecimento de populações nas montanhas da região equatorial africana levou à seguinte distribuição em função da altitude e da idade geológica das cadeias montanhosas:
| Kilimanjaro Dendrosenecio kilimanjari D. johnstonii | |||
| Cordilheira de Aberdare D. battiscombei D. brassiciformis |
Monte Meru (Tanzânia) D. meruensis | ||
| Monte Kenya D. keniodendron D. keniensis D. battiscombei |
Colinas Cherangani D. cheranganiensis |
Monte Elgon D. elgonensis |
|
| Cordilheira de Aberdare D. keniodendron |
Montanhas Virunga D. erici-rosenii | ||
| Montanhas Mitumba D. erici-rosenii | |||
| Montanhas Ruwenzori D. erici-rosenii D. adnivalis | |||
A interpretação biogeográfica da filogenia molecular sugere que no milhão de anos mais recente, os primeiros senécios gigantes estabeleceram-se nas altitudes mais elevadas do Monte Kilimanjaro e tornaram-se na espécie Dendrosenecio kilimanjari.[3]
Ao progressivamente se estabelecerem em altitudes inferiores nas encostas da montanha, adaptando-se ao ambiente diferente das altitudes mais baixas do Monte Kilimanjaro, transformaram-se numa nova espécie, a Dendrosenecio johnstonii.
Algumas sementes encontraram um caminho para o Monte Meru e estabeleceram-se como a espécie Dendrosenecio meruensis, outras encontraram um caminho para ir do Monte Kilimanjaro para a Cordilheira de Aberdare e estabeleceram-se como Dendrosenecio battiscombei. A migração da D. battiscombei para o habitat alpino húmido de Aberdares resultou na formação da espécie Dendrosenecio brassiciformis.[3]
A dispersão dos Aberdares para o Monte Quénia estabeleceu uma segunda população isolada de D. battiscombei. A especiação altitudinal no Monte Quénia resultou na formação das espécies Dendrosenecio keniodendron e Dendrosenecio keniensis, esta última do tipo anão. A dispersão do Monte Quénia de volta para os Aberdares estabeleceu uma segunda população insular de D. keniodendron.[3]
A dispersão dos Aberdares para as colinas de Cherangani deu origem a duas subespécies de D. cheranganiensis: (1) D. cheranganiensis subsp. cheranganiensis; (2) e a (sub)especiação altitudinal para o habitat alpino deu origem a D. cheranganiensis subsp. dalei. A dispersão dos Aberdares para o Monte Elgon deu origem a D. elgonensis, que é um ponto onde várias subespécies divergem e se dispersam: do Monte Elgon para as Montanhas Virunga deu origem a D. erici-rosenii; do Monte Elgon para o Monte Kahuzi (Montanhas Mitumba) deu origem a uma segunda população de D. erici-rosenii e a dispersão das Montanhas Virunga para as Montanhas Ruwenzori deu origem a uma terceira população.[3]
Paralelismo evolutivo
As comunidades encontradas nestas montanhas africanas de Dendrosenecio e de Lobelia com espécies de grande dimensões quando comparadas com os géneros filogeneticamente próximos, são um exemplo excecional de evolução paralela (ou evolução convergente) a ocorrer de forma repetida entre estes dois grupos. A existência destas espécias gigantes fornece provas de que as caraterísticas invulgares destas plantas são uma resposta evolutiva a um habitat desafiante e a um ambiente natural que pode ser facilmente descrito para análise biogeográfica.[3]
Uniformidade citológica
Foi encontrada pouca variação na filogenia molecular entre as 40 colecções de senécio gigante registadas (40 acessos), mas como grupo diferem significativamente de Cineraria deltoidea, o parente mais próximo conhecido.[8]
O número cromossómico do gametófito (o número de cromossomas em cada célula) no género gigante Dendrosenecio é n = 50, e para as lobélias gigantes, especificamente para o subgénero Lobelia tupa, da secção Rhynchopetalum das Lobeliaceae, é n = 14.[16]
Apenas 5 das 11 espécies de senécio gigante e três das 21 espécies de lobélia gigante da África oriental continuam por analisar. Apesar de ambos os grupos serem poliploides, presume-se que o género Dendrosenecio seja decaploide (dez conjuntos; 10x) e o género Lobelia tetraploide (quatro conjuntos; 4x), sendo que as suas radiações adaptativas não envolveram alterações subsequentes no número de cromossomas.[16]
A uniformidade citológica dentro de cada grupo, embora forneça evidências circunstanciais de que descendem de um único ancestral comum e simplifique as interpretações de análise cladística, não fornece suporte positivo nem negativo para um possível papel da poliploidia na evolução da forma de crescimento gigante em roseta.[16]
Referências
- ↑ a b c «Index Nominum Genericorum database». International Code of Botanical Nomenclature. Smithsonian Institution. 1978. Consultado em 4 de maio de 2008
- ↑ Botanic Garden; Botanical Museum Berlin-Dahlem (1978). «Entry for Dendrosenecio». Names in Current Use for Extant Plant Genera. Freie Universität Berlin. Consultado em 4 de maio de 2008
- ↑ a b c d e f g h i j k l m n Knox, Eric B. (2004). «Adaptive radiation of African montane plants». In: Ulf Dieckmann; Michael Doebeli; Diethard Tautz; Johan A. J. Metz. Adaptive Speciation. [S.l.]: Cambridge University Press. 476 páginas. ISBN 0-521-82842-2. Consultado em 29 de março de 2008
- ↑ «Ultra-prominências de África: 84 Montanhas com proeminência de 1 500 m ou superior». Peaklist. Consultado em 5 de maio de 2008
- ↑ «Dendrosenecio B. Nord». Tropicos. Consultado em 21 de julho de 2019
- ↑ Nordenstam, Rune Bertil. 1978. Opera Botanica 44: 40
- ↑ Tropicos, Dendrosenecio B. Nord.
- ↑ a b c d e Knox, Eric B.; Jeffrey D. Palmer (24 de outubro de 1995). «Chloroplast DNA variation and the recent radiation of the giant senecios (Asteraceae) on the tall mountains of eastern Africa». National Academy of Sciences. Proceedings of the National Academy of Sciences. 92 (22): 10349–10354. Bibcode:1995PNAS...9210349K. PMC 40794
 . PMID 7479782. doi:10.1073/pnas.92.22.10349
. PMID 7479782. doi:10.1073/pnas.92.22.10349
- ↑ Roosevelt, Theodore; Edmund Heller (18 de setembro de 2007). «Logically». Life-histories of African Game Animals 1 ed. [S.l.]: C. Scribner's Sons. ISBN 978-1-4446-8030-0. Consultado em 28 de março de 2008.
Groundsels of several species are found throughout the world as common roadside weeds, but nowhere except in the highlands of Africa do they exhibit such large tree forms.
- ↑ a b Bussmann, Fainer W. (Junho de 2006). «Vegetation zonation and nomenclature of African Mountains - An overview». Lyonia. Consultado em 27 de abril de 2008
- ↑ a b Weischet, Wolfgang; Endlicher, Wilfried (2000). Regionale Klimatologie Teil 2 Die Alte Welt: Europa - Afrika - Asien (Regional climatology, Part 2: The old world: Europe - Africa - Asia). [S.l.]: Borntraeger. p. 625. ISBN 978-3-443-07119-6
- ↑ Hedberg, Olov (1955). «Vegetation belts of the East-African mountains». Proceedings of the Linnean Society of London (Botany). 165: 134–136. doi:10.1111/j.1095-8312.1955.tb00730.x
- ↑ Hedberg, Karl Olov (1964). «Features of afroalpine plant ecology». Acta Phytogeographica Suecica. 49: 1–144. ISBN 91-7210-049-4. Consultado em 4 de maio de 2008
- ↑ Hauman, L. L. (1955). «La 'region afroapline' en phytogeographie centro africaine». Webbia. 11: 467–489. doi:10.1080/00837792.1956.10669644
- ↑ Hedberg, Olov (1951). «Vegetation belts of the East-African mountains.». Svensk Botanisk Tidskrift. 45: 141–196
- ↑ a b c Knox, Eric B.; Robert R. Kowal (Julho de 1993). «Chromosome Numbers of the East African Giant Senecios and Giant Lobelias and their Evolutionary Significance» (PDF). Botanical Society of America. American Journal of Botany. 80 (7): 847–853. JSTOR 2445604. doi:10.2307/2445604. hdl:2027.42/141794
Ligações externas
- Cyrille Chatelain; Laurent Gautier; Raoul Palese. «Dendrosenecio (Hauman ex Hedberg) B.Nord.». African Flowering Plants Database. Conservatoire et Jardin botaniques de la Ville de Genève. Consultado em 29 de março de 2008 [ligação inativa]
- Germplasm Resources Information Network (GRIN) (29 de janeiro de 2005). «Genus: Dendrosenecio (Hauman ex Hedberg) B. Nord.». Taxonomy for Plants. USDA, ARS, National Genetic Resources Program, National Germplasm Resources Laboratory, Beltsville, Maryland. Consultado em 29 de março de 2008. Cópia arquivada em 28 de maio de 2010