Cebus imitator
Cebus imitator
| |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
2.jpg) | |||||||||||||||||||
| Estado de conservação | |||||||||||||||||||
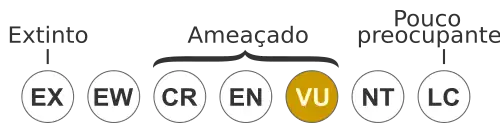 Vulnerável [1] | |||||||||||||||||||
| Classificação científica | |||||||||||||||||||
| |||||||||||||||||||
| Distribuição geográfica | |||||||||||||||||||
![Distribuição de Cebus imitator e C. capucinus.[2] C. imitator cobre a porção centro-americana da distribuição, exceto a parte mais oriental do Panamá.](./_assets_/0c70a452f799bfe840676ee341124611/Cebus_capucinus_and_imitator_Range_Map.png) Distribuição de Cebus imitator e C. capucinus.[2] C. imitator cobre a porção centro-americana da distribuição, exceto a parte mais oriental do Panamá.
| |||||||||||||||||||
A espécie Cebus imitator, que é popularmente chamada de "Panamanian white-faced capuchin" no inglês (capuchinho-de-cara-branca-panamenho em tradução livre), é um macaquinho do Novo Mundo de tamanho médio da família Cebidae, subfamília Cebinae. Nativo das florestas da América Central, o capuchinho-de-cara-branca é importante para a ecologia da floresta tropical por seu papel na dispersão de sementes e de pólen.
Nos últimos anos, a espécie ganhou popularidade na mídia americana, particularmente na série de filmes Piratas do Caribe. É um macaco altamente inteligente e foi treinado para auxiliar pessoas paraplégicas. É um macaco de tamanho médio, pesando até 3.9 kg. É predominantemente preto, mas com rosto rosado e branco em grande parte da frente do corpo, o que lhe confere seu nome comum. Possui uma cauda preênsil distinta, frequentemente carregada enrolada e usada para apoiar o macaco quando se alimenta sob um galho.
Na natureza, o C. imitator é versátil, vivendo em diversos tipos de floresta e se alimentando de vários tipos de comida, incluindo frutas, outros materiais vegetais, invertebrados e pequenos vertebrados. Vive em bandos que podem exceder 20 indivíduos e incluem machos e fêmeas. É notado por seu uso de ferramentas, incluindo esfregar plantas no corpo em um aparente uso de medicina herbal e usar ferramentas como armas ou para acessar alimentos. É um macaco de longa vida, com uma idade máxima registrada de mais de 54 anos.
Os capuchinhos-de-cara-branca-panamenhos são altamente sociais, vivendo em grupos de, em média, 16 indivíduos, cerca de três quartos dos quais são fêmeas. Os grupos consistem em fêmeas relacionadas, machos imigrantes e seus filhotes. Em média, as fêmeas dão à luz a cada 27 meses, embora acasalem durante todo o ano. As fêmeas tendem a permanecer no grupo original, enquanto os machos deixam o grupo natal aos quatro anos e mudam de grupo a cada quatro anos depois disso. Machos e fêmeas exibem diferentes comportamentos de dominância dentro do grupo.
Taxonomia

.jpg)
C. imitator é um membro da família Cebidae, que abrange macacos-prego e macacos-de-cheiro. Até o século XXI, a espécie era considerada coespecífica com C. capucinus, o macaco-prego-de-cara-branca (também conhecido como "capuchinho-de-cara-branca-colombiano), mas como uma subespécie distinta, C. capucinus imitator.[3] Alguns primatólogos continuam a considerar os capuchinhos-de-cara-branca panamenhos e colombianos como uma única espécie.[4] É um membro do grupo de espécies C. capucinus dentro do gênero Cebus, que também inclui o capuchinho-de-cara-branca-colombiano, o caiarara e o macaco-prego-kaapori.[3] Este gênero também é referido como "capuchinhos gráceis".
Em 2012, um estudo de Boubli e colaboradores demonstrou que C. imitator e C. capucinus divergiram há 2 milhões de anos.[5][6] O estudo de Boubli também indicou que os capuchinhos-de-cara-branca de Honduras, anteriormente considerados uma possível subespécie distinta, C. capucinus limitaneus, não eram geneticamente distintos de C. imitator.[5]
C. imitator é a espécie de macaco-prego mais estudada.[6] Embora muitos estudos anteriores tenham sido realizados usando o nome científico C. capucinus, até 2014 não havia estudos de campo do capuchinho-de-cara-branca-colombiano, então todos esses estudos referiam-se ao C. imitator.[6]
Descrição física
Como outros macacos do gênero Cebus, C. imitator é nomeada em homenagem à ordem dos frades capuchinhos – os capuzes desses frades lembram muito a coloração da cabeça do macaco.[7][8] A espécie apresenta pelagem predominantemente preta, com pelos brancos a amarelados no pescoço, garganta, peito, ombros e braços superiores.[9] O rosto é rosado ou de cor branca-creme e pode ter marcas identificadoras, como sobrancelhas escuras ou manchas de pelagem escura.[9][10][11] Uma área de pelagem preta na coroa da cabeça é distinta.[9][12] Possui uma cauda preênsil frequentemente mantida enrolada, dando aos capuchinhos-de-cara-branca o apelido de "cauda-de-anel".[9][13]
Os adultos alcançam um comprimento entre 335 e 453 mm, excluindo a cauda, e um peso de até 3.9 kg.[9][12] A cauda é mais longa que o corpo, com até 551 mm de comprimento.[9][12] Os machos são cerca de 27% maiores que as fêmeas.[14] O cérebro de um capuchinho-de-cara-branca tem cerca de 79.2 g, que é maior que o de várias espécies de macacos maiores, como o bugio Alouatta palliata.[12][15]
C. imitator é semelhante ao capuchinho-de-cara-branca-colombiano na aparência, exceto que as fêmeas panamenhas têm tufos frontais alongados marrons ou acinzentados, que contrastam com as bochechas e garganta brancas puras.[4][16]
Comportamento
Estrutura social

C. imitator é um animal diurno e arborícola.[9] No entanto, desce ao chão com mais frequência que muitos outros macacos do Novo Mundo.[17] Move-se principalmente caminhando sobre os quatro membros.[18] Vive em bandos, ou grupos, de até 40 indivíduos (média de 16, faixa de 4–40)[19] e tem uma proporção de sexo adulto macho/fêmea de 0,71 em média (faixa de 0,54–0,88).[19] Com raras exceções, as fêmeas passam toda a vida com suas parentes fêmeas.[19][20][21]
Os machos migram para novos grupos sociais várias vezes durante a vida, migrando pela primeira vez entre 20 meses e 11 anos de idade.[22][23] A idade mediana de migração na população de Santa Rosa é de 4,5 anos.[22] Os machos às vezes migram sozinhos, mas mais frequentemente migram na companhia de outros machos que muitas vezes são seus parentes.[21][22][23] Uma das características incomuns da estrutura de parentesco de C. imitator, em relação a outras espécies de primatas, é o alto grau de parentesco dentro dos grupos, resultado dos longos mandatos dos machos alfa que geram a maioria dos filhotes.[24][25] Os machos alfa são conhecidos por manter suas posições por até 17 anos nesta espécie[24][25] e isso os coloca na posição incomum de estarem disponíveis para gerar filhotes de suas filhas e netas, que produzem seus primeiros filhotes por volta dos 6–7 anos de idade.[19][25] Tipicamente, no entanto, os machos alfa não se reproduzem com suas próprias filhas, embora gerem virtualmente todos os filhotes produzidos por fêmeas não relacionadas a eles.[24] Os machos subordinados que são aliados do macho alfa na defesa do grupo são os que geram os filhotes das filhas do macho alfa. O alto grau de monopolização de acasalamentos pelos machos alfa resulta em um número excepcionalmente grande de meio-irmãos paternos e irmãos completos nesta espécie em relação a outras espécies de primatas.[25]
O parentesco é um fator organizador importante na estruturação das relações sociais entre fêmeas.[25] Particularmente em grupos maiores, as fêmeas preferencialmente associam-se, cuidam e fornecem apoio de coalizão às suas parentes matrilineares. Elas não exibem uma preferência semelhante por suas meias-irmãs paternas, o que pode significar que só são capazes de reconhecer parentesco pela linha materna.[25] A hierarquia de dominância também é um fator organizador importante, com fêmeas frequentemente cuidando e associando-se a fêmeas mais próximas a elas na hierarquia de dominância.[25] As duplas fêmea-fêmea cuidam muito mais do que as duplas macho-fêmea e macho-macho.[26] A agressão de coalizão é comum tanto entre machos quanto entre fêmeas, e os capuchinhos parecem ter uma excelente compreensão da estrutura de alianças em seu grupo. Por exemplo, quando lutam, eles sensatamente recrutam ajuda de alguém que é tanto mais alto em hierarquia quanto mais amigo deles do que de seu oponente.[27]
As fêmeas capuchinhas têm hierarquias de dominância lineares.[26][28] Em contraste com muitos macacos do Velho Mundo, como Macaca, em que as fêmeas herdam socialmente a posição logo abaixo de suas mães e acima de suas irmãs mais velhas, os capuchinhos não têm uma classificação altamente previsível dentro de suas matrilinhagens.[25] Os machos são tipicamente dominantes sobre as fêmeas.[29] O macho alfa é sempre fácil de discernir, mas às vezes há classificações ambíguas entre machos subordinados.[21][30] As relações macho-macho são tensas, e a afiliação entre machos é tipicamente expressa por descanso em contato, brincadeiras ou sexo não-concepcional [en] em vez de cuidado.[30][31] Os machos cooperam em coalizões contra predadores potenciais e também na defesa do grupo contra outros machos.[21][30][32] Ocasionalmente, a agressão de coalizão masculina torna-se tão violenta que machos são mortos, particularmente se encontrados vagando pela floresta sem a companhia de aliados.[21][33] Como a agressão de outros machos capuchinhos é a principal causa de morte (além da caça furtiva por humanos, onde há contato entre humanos e capuchinhos), aliados machos são cruciais para autodefesa durante a migração e para ajudar na tomada de outros grupos.[21] A emigração masculina para um novo bando ocorre tipicamente a cada 4 anos, então a maioria dos machos está em constante perigo de ter que se defender contra outros grupos de machos.[8][34][35][36]
Machos imigrantes frequentemente matam filhotes jovens quando assumem um grupo.[21][37][38] As fêmeas se unem para defender seus filhotes de machos infanticidas, mas raramente conseguem salvar seus filhotes.[21] Como os filhotes inibem suas mães de ovular ao amamentar com frequência, os machos conseguem trazer as fêmeas ao estro mais cedo ao matar os filhotes e, assim, interromper a amamentação; isso tem o efeito de aumentar suas oportunidades de reprodução.[21][38][39] As fêmeas frequentemente acasalam com os assassinos de seus filhotes e, com o tempo, geralmente se tornam tão solidárias ao novo macho alfa quanto eram ao anterior.[21] O macho alfa ajuda a defender as fêmeas de machos subordinados dentro do grupo, bem como de machos infanticidas de outros grupos.[21][29]
Interações entre grupos
Os bandos de capuchinhos-de-cara-branca-panamenhos ocupam áreas de vida de 32 a 86 ha.[12] Eles viajam entre 1 e 3 km diariamente, com média de 2 km por dia.[40] Embora se engajem em atividades descritas como "territoriais", pesquisas mais recentes indicam que os bandos de capuchinhos-de-cara-branca tendem a se comportar agressivamente com outros bandos de capuchinhos-de-cara-branca, independentemente de onde se encontram, e a agressão não é necessariamente destinada a excluir outros bandos de uma área de vida específica.[41]
As áreas de vida se sobrepõem extensivamente,[32][42] então os grupos não são territoriais no sentido mais estrito da palavra. Talvez devido à intensidade da competição macho-macho e à ameaça de infanticídio, as interações entre grupos são tipicamente hostis: os machos exibem-se agressivamente uns contra os outros e às vezes se envolvem em agressão física (até matando um oponente), enquanto as fêmeas pegam seus filhotes e fogem.[32][33] Tipicamente, os machos são os principais participantes em encontros agressivos entre grupos, e parece provável que os machos estejam defendendo o acesso às fêmeas em seus grupos.[32] Machos alfa, que têm o maior interesse reprodutivo no grupo, participam em uma taxa mais alta que machos subordinados.[32] Grupos com mais machos têm uma vantagem sobre grupos com menos machos, mas a localização do encontro dentro da área de vida também importa; grupos menores derrotam grupos maiores quando o confronto ocorre na área central da área de vida do grupo menor.[42]
Interações interespecíficas
C. imitator às vezes interage com outras espécies simpátricas de macacos. Os capuchinhos-de-cara-branca-panamenhos às vezes viajam com e até cuidam de macacos-aranha-de-Geoffroy.[12][40] No entanto, interações agressivas entre os capuchinhos e os macacos-aranha também ocorrem.[43] Interações entre C. imitator e o bugio A. palliata são pouco frequentes e às vezes resultam nos capuchinhos ameaçando os bugios maiores.[40] No entanto, associações afiliativas entre os capuchinhos e bugios ocorrem às vezes, principalmente envolvendo juvenis brincando juntos.[43]
Embora as espécies de capuchinhos sul-americanos frequentemente viajem e se alimentem junto com macacos-de-cheiro, C. imitator raramente se associa com o Saimiri oerstedii. Isso parece estar relacionado à distribuição mais irregular e dispersa de recursos alimentares na América Central e ao fato de haver menos sobreposição alimentar entre o S. oerstedii e o capuchinho-de-cara-branca do que entre seus equivalentes sul-americanos. Portanto, há menos benefício para o macaco-de-cheiro em se associar ao C. imitator para explorar o conhecimento da distribuição de recursos alimentares. Além disso, em comparação com seus equivalentes sul-americanos, os machos capuchinhos-de-cara-branca-panamenhos estão relativamente mais alertas a machos rivais do que a predadores, reduzindo os benefícios de detecção de predadores que o macaco-de-cheiro S. oerstedii recebe ao se associar ao C. imitator em comparação com seus equivalentes sul-americanos. Como os macacos-de-cheiro geralmente iniciam interações com os capuchinhos na América do Sul, o fato de associações semelhantes imporiam custos de forrageamento mais altos e trariam menos benefícios de detecção de predadores ao macaco-de-cheiro leva a menos associações com o C. imitator.[14][44][45][46]
Na ilha de Jicarón [en], os capuchinhos-de-cara-branca-panamenhos coexistem com bugio-da-ilha-Coiba (A. coibensis coibensis).[47] No início de 2022, câmeras ativadas por movimento operando desde 2017 em Jicarón para observar o uso de ferramentas pelos capuchinhos detectaram, pela primeira vez, machos capuchinhos carregando filhotes de bugios nas costas enquanto caminhavam ou esmagavam sementes com pedras. Pesquisas adicionais indicaram que, durante 2022 e 2023, pelo menos cinco machos capuchinhos sequestraram pelo menos 11 filhotes de bugios e os carregaram nas costas. Embora os capuchinhos não tenham machucado os filhotes de bugios, eles foram incapazes de cuidar ou alimentá-los adequadamente, e pelo menos quatro — e provavelmente a maioria ou todos — dos filhotes de bugios morreram. Quando anunciaram suas descobertas em maio de 2025, os pesquisadores concluíram que as câmeras capturaram um comportamento que surgiu recentemente como uma "moda" entre uma única população de machos capuchinhos, e descreveram o comportamento dos capuchinhos como "a primeira documentação conhecida de uma tradição social em que animais sequestram e carregam repetidamente filhotes de outra espécie — sem qualquer benefício claro para si mesmos."[48][49][50]
Várias espécies de animais não primatas tendem a seguir bandos de macacos-de-cara-branca ou são de outra forma atraídas por sua presença. Queixadas e cutias são atraídas por capuchinhos-de-cara-branca que se alimentam, procurando frutas que os capuchinhos deixam cair.[40] Várias espécies de aves também são conhecidas por seguir capuchinhos-de-cara-branca-panamenhos em busca de comida. Estas incluem o gavião-ripina, o gavião-branco e o gavião-miúdo.[40]
Dieta

C. imitator é uma espécie onívora. Seus alimentos primários são frutas e insetos.[8] Ele forrageia em todos os níveis da floresta, incluindo o chão.[40] Métodos para encontrar comida incluem arrancar cascas de árvores, procurar em folhagem, quebrar galhos de árvores mortas, virar pedras e usar pedras como bigornas para quebrar frutas duras.[51] Sua cauda preênsil auxilia na alimentação, ajudando a suportar o macaco quando forrageia abaixo dos galhos.[40]
Frutas podem compor entre 50% e 67% ou mais da dieta do capuchinho.[8] Em um estudo no Panamá, capuchinhos-de-cara-branca comeram 95 espécies diferentes de frutas.[8] Entre suas frutas favoritas estão figos da família Moraceae, mangas e frutas relacionadas da família Anacardiaceae, frutos tipo feijão da família Leguminosae e frutas da família Rubiaceae.[52] Ele também consome frutas de Euphorbiaceae, como o feijão-saltador mexicano Sebastiania pavoniana.[53]

Geralmente, só come frutas maduras, testando a maturidade pelo cheiro, sabor e toque.[8] Normalmente, come apenas a polpa e o suco, cuspindo as sementes e fibras.[8] Outros materiais vegetais consumidos incluem flores, folhas jovens, sementes de certas plantas e bromélias.[8][54] Ele também usa as bromélias como fonte de água, bebendo a água que fica presa dentro delas.[8] No Parque Nacional de Carara [en], os capuchinhos têm uma dieta variada, além do acima, de frutas e flores de banana, sementes de helicônia, frutos de Tabernaemontana e caules de Anacardiaceae.[55]
Presas de insetos consumidas incluem larvas de besouro, lagartas de borboletas e mariposas, formigas, vespas e larvas de formigas e vespas.[8] Ele também consome presas maiores, como aves, ovos de aves, sapos, lagartos, caranguejos, moluscos e pequenos mamíferos.[8][56] A população na Guanacaste, Costa Rica, é particularmente notada por caçar esquilos, pica-paus, maitacas-de-testa-branca[8] e filhotes de quati.[57] A quantidade de presas vertebrados consumidas varia por bando.[8] Mesmo bandos vizinhos podem mostrar diferenças significativas em suas dietas.[51]
A dieta pode variar entre a estação chuvosa e a seca. Por exemplo, em Guanacaste, Costa Rica, C. imitator pode comer uma ampla variedade de frutas, bem como lagartas, na estação chuvosa inicial (junho a novembro).[52] Mas durante a estação seca, apenas figos e algumas outras frutas estão disponíveis.[52] Então, durante a estação seca, insetos quitinosos, larvas de formigas e vespas e vertebrados tornam-se particularmente importantes na dieta da espécie.[52] O acesso à água também pode se tornar um problema durante a estação seca. C. imitator gosta de beber diariamente, então em florestas onde os poços de água secam durante a estação seca, pode haver competição entre bandos pelo acesso aos poços de água remanescentes.[52]
Uso de ferramentas
Os capuchinhos são considerados entre os mais inteligentes dos macacos do Novo Mundo; eles foram objeto de muitos estudos sobre comportamento e inteligência. A inteligência dos capuchinhos é considerada uma adaptação para apoiar seus hábitos alimentares; eles dependem de fontes de alimentos efêmeras que podem ser difíceis de encontrar. Em um estudo particular conduzido em 2007, os capuchinhos foram classificados entre os dez primatas mais inteligentes, ficando em segundo lugar entre os macacos do Novo Mundo, atrás dos macaco-aranha.[58]
O uso de ferramentas de pedra é uma diferença marcante entre os capuchinhos gráceis do gênero Cebus e os capuchinhos robustos do gênero Sapajus. Embora generalizado em capuchinhos robustos, apenas um caso de uso habitual de ferramentas de pedra foi relatado por capuchinhos gráceis. Uma população de capuchinhos-de-cara-branca-panamenhos encontrada no Parque Nacional de Coiba no Panamá foi observada usando martelo de pedra e bigornas para processar frutas de castanheiras, Bactris major e coqueiros e invertebrados como caracóis Nerita, caranguejos-eremitas e outros caranguejos.[59][60]
C. imitator é conhecido por esfregar partes de certas plantas em seus pelos. Plantas usadas dessa maneira incluem frutas cítricas, trepadeiras dos gêneros Piper e Clematis, pente-de-macaco (gênero Sloanea), comigo-ninguém-pode e anonas.[8][56] Formigas e milípedes também são usados dessa maneira.[8] Não se sabe definitivamente para que serve esse esfregamento, mas pode deter parasitas como carrapatos e insetos, ou pode funcionar como fungicida, bactericida ou agente anti-inflamatório.[8] Alternativamente, pode ser uma forma de marcação de odor.[8] C. imitator também usa ferramentas de outras maneiras. Ele foi conhecido por bater em cobras com varas para se proteger ou para fazer a cobra soltar um filhote,[8] e às vezes usa varas como sondas para explorar aberturas.[61] Em cativeiro, foi conhecido por usar ferramentas para alcançar comida ou para se defender, e em um caso, um capuchinho-de-cara-branca usou um macaco-de-cheiro como projétil, arremessando-o contra um observador humano.[8]
A inteligência e a capacidade de usar ferramentas da espécie permitem que ele seja treinado para ajudar paraplégicos.[62] Outras espécies de macaco-prego também são treinadas dessa maneira.[63] Capuchinhos-de-cara-branca-panamenhos também podem ser treinados para papéis em televisão e cinema, como Marcel na série de televisão Friends.[64] Eles também foram tradicionalmente usados como macacos de tocadores de realejo.[65]
Comunicação

C. imitator é barulhento.[9] Chamados altos, como latidos e tosses, são usados para comunicar avisos de ameaça, e chamados mais suaves, como guinchos, são usados em discursos íntimos.[8] Diferentes tipos de ameaças, como uma ameaça de um animal terrestre versus uma ameaça de uma ave, invocam vocalizações diferentes.[40] Expressões faciais e odor também são importantes para a comunicação.[66] Ele às vezes se engaja em uma prática conhecida como "lavagem de urina", em que o macaco esfrega urina em seus pés.[67] O propósito exato dessa prática é desconhecido, mas pode ser uma forma de sinal olfativo.[66]
Reprodução

C. imitator usa um sistema de acasalamento poligâmico em que um macho pode acasalar com várias fêmeas.[40] Embora o macho dominante não monopolize a reprodução, estudos mostraram que o macho dominante tende a ser o pai da maioria dos filhotes.[35] Embora uma fêmea possa acasalar com vários machos, o macho dominante pode ser mais propenso a copular quando a fêmea está no pico de fertilidade.[35][68] No entanto, há evidências de que machos dominantes evitam se reproduzir com suas próprias filhas que são membros do bando.[69] Tal evitação é rara entre primatas do Novo Mundo.[69]
A cópula leva cerca de 2 minutos, e o período de gestação é de 5 a 6 meses.[40] Normalmente, nasce um único filhote, mas gêmeos ocorrem ocasionalmente. A maioria dos nascimentos ocorre durante a estação seca, de dezembro a abril.[12][40] O filhote é carregado nas costas da mãe por cerca de 6 semanas.[40] Após cerca de 4 a 5 semanas, ele pode se afastar da mãe por breves períodos e, por volta dos 3 meses, pode se mover independentemente, embora alguns filhotes sejam majoritariamente independentes mais cedo. O desmame ocorre entre 6 e 12 meses. Enquanto a mãe descansa, o jovem passa a maior parte do tempo forrageando ou brincando, seja sozinho ou com outros juvenis.[40] Os capuchinhos praticam altos níveis de aloparentalidade, em que macacos além da mãe ajudam a cuidar do filhote.[70] Os filhotes são carregados por aloparentes com mais frequência entre 4 e 6 semanas de idade.[21] Machos e fêmeas participam da aloparentalidade.[21][40]
Como outras espécies de capuchinhos, C. imitator amadurece lentamente. A maturidade sexual pode ser alcançada aos 3 anos.[65] Mas, em média, as fêmeas dão à luz pela primeira vez aos 7 anos e dão à luz a cada 26 meses depois disso.[14] Os machos atingem a maturidade reprodutiva aos 10 anos.[14] C. imitator tem uma longa expectativa de vida para seu tamanho. A expectativa de vida máxima registrada em cativeiro é de mais de 54 anos.[14]
Distribuição e habitat
C. imitator é encontrada em grande parte da América Central. Sua distribuição inclui grande parte de Honduras, Nicarágua, Costa Rica e Panamá.[4][2] Também foi relatado ocorrer no leste da Guatemala e no sul de Belize, mas esses relatos não estão confirmados.[2] Está entre os macacos mais comumente vistos nos parques nacionais da América Central, como Parque Nacional Manuel Antonio, Parque Nacional Corcovado, Parque Nacional de Santa Rosa e Parque Nacional Soberanía.[71] Aparece no verso da nota de 5.000 colones da Costa Rica.
Embora o capuchinho-de-cara-branca seja muito comum na Costa Rica e no Panamá, o macaco foi amplamente extirpado de Honduras e grande parte da Nicarágua. Muitos capuchinhos hondurenhos foram capturados e realocados para a ilha de Roatán, e muitos capuchinhos nicaraguenses foram capturados e realocados para a ilha de Ometepe. Na Nicarágua, capuchinhos selvagens ainda podem ser facilmente avistados em regiões ao redor de Masaya, bem como ao redor de Bluefields e outras localizações ao longo da costa sul do Caribe. Eles são vistos, na natureza, diariamente por visitantes que escalam um dos vulcões na Ilha de Ometepe.[72]
É encontrado em muitos tipos diferentes de floresta, incluindo florestas maduras e secundárias, florestas perenes e decíduas, secas e úmidas, além de mangues e florestas montanas.[9][65] No entanto, parece preferir florestas primárias ou secundárias avançadas.[40] Além disso, uma maior densidade de capuchinhos-de-cara-branca é encontrada em áreas de floresta mais antigas e em áreas contendo florestas perenes, bem como em áreas com maior disponibilidade de água durante a estação seca.[73]
Estado de Conservação

C. imitator é considerado vulnerável do ponto de vista da conservação pela IUCN.[1] Está ameaçado por desmatamento, caça para o comércio de animais de estimação e, às vezes, para carne de caça, além de ataques por parte de agricultores que os consideram uma ameaça potencial.[1] No entanto, o desmatamento pode impactar seu principal predador, o gavião-real, mais do que afeta diretamente C. imitator, de modo que, no saldo líquido, o desmatamento pode não ser tão prejudicial ao status do capuchinho.[8]
A espécie pode se adaptar à fragmentação florestal melhor que outras espécies devido à sua capacidade de viver em uma ampla variedade de tipos de floresta e explorar diversas fontes de alimento.[74] C. imitator é importante para seus ecossistemas como dispersor de sementes e pólen.[8][65] Ele também impacta o ecossistema ao comer insetos que atuam como pragas para certas árvores, ao podar certas árvores, como Gustavia superba [en] e Bursera simaruba, fazendo com que gerem mais ramos e possivelmente frutos adicionais, e ao acelerar a germinação de certas sementes ao passarem pelo trato digestivo do capuchinho.[8] Além disso, C. imitator às vezes mata plantas de Vachellia collinsii quando rasga seus ramos para acessar colônias de formigas residentes.[8]
Referências
- ↑ a b c Williams-Guillén, K.; Rosales-Meda, M.; Méndez-Carvajal, P.G.; Solano-Rojas, D.; Urbani, B; Lynch-Alfaro, J.W. (2021). «Cebus imitator». Lista Vermelha de Espécies Ameaçadas. 2021: e.T81265980A191708420. doi:10.2305/IUCN.UK.2021-1.RLTS.T81265980A191708420.en
 . Consultado em 29 de novembro de 2021
. Consultado em 29 de novembro de 2021
- ↑ a b c Rylands, A.; Groves, C.; Mittermeier, R.; Cortes-Ortiz, L.; Hines, J. (2006). «Taxonomy and Distributions of Mesoamerican Primates». In: Estrada, A.; Garber, P.; Pavelka, M.; Luecke, L. New Perspectives in the Study of Mesoamerican Primates. New York: Springer. pp. 40–43. ISBN 978-0-387-25854-6
- ↑ a b Groves, C.P. (2005). Wilson, D. E.; Reeder, D. M, eds. Mammal Species of the World 3.ª ed. Baltimore: Johns Hopkins University Press. p. Cebus. ISBN 978-0-8018-8221-0. OCLC 62265494
- ↑ a b c Mittermeier, Russell A.; Rylands, Anthony B. (2013). Mittermeier, Russell A.; Rylands, Anthony B.; Wilson, Don E., eds. Handbook of the Mammals of the World: Volume 3, Primates. [S.l.]: Lynx. pp. 412–413. ISBN 978-8496553897
- ↑ a b Boubli, Jean P.; et al. (2012). «Cebus Phylogenetic Relationships: A Preliminary Reassessment of the Diversity of the Untufted Capuchin Monkeys» (PDF). American Journal of Primatology. 74 (4): 1–13. PMID 22311697. doi:10.1002/ajp.21998. Consultado em 30 de dezembro de 2018. Arquivado do original (PDF) em 4 de março de 2016
- ↑ a b c Lynch Alfaro, Jessica; et al. (2014). «Capuchin Monkey Research Priorities and Urgent Issues» (PDF). American Journal of Primatology. 76 (8): 1–16. PMID 24668460. doi:10.1002/ajp.22269. Consultado em 30 de dezembro de 2018. Arquivado do original (PDF) em 10 de agosto de 2017
- ↑ «Capuchin Franciscans F.A.Q.». Capuchin Franciscans Vocation Office Province of Saint Joseph. Consultado em 1 de setembro de 2008. Arquivado do original em 25 de julho de 2011
- ↑ a b c d e f g h i j k l m n o p q r s t u v w x Wainwright, M. (2002). The Natural History of Costa Rican Mammals. [S.l.]: Zona Tropical. pp. 135–139. ISBN 978-0-9705678-1-9
- ↑ a b c d e f g h i Emmons, L. (1997). Neotropical Rainforest Mammals A Field Guide Second ed. [S.l.]: University of Chicago Press. pp. 130–131. ISBN 978-0-226-20721-6
- ↑ Luedtke, Karen (2012). Jungle Living: A look at life and social behavior of man and monkey in Central American. [S.l.: s.n.] pp. 40–45. ISBN 978-0-9832448-2-0
- ↑ Luedtke, K. (2012). Jungle Living: A look at life and social behavior of man and monkey in central america. [S.l.: s.n.] p. 45. ISBN 978-0-9832448-2-0
- ↑ a b c d e f g Rowe, N. (1996). The Pictorial Guide to the Living Primates. [S.l.]: Pogonias Press. p. 95. ISBN 978-0-9648825-0-8
- ↑ «Medical Dictionary Capuchin Monkey». Farlex Inc. Consultado em 1 de setembro de 2008
- ↑ a b c d e Jack, K. (2007). «The Cebines». In: Campbell, C.; Fuentes, A.; MacKinnon, K.; Panger, M.; Bearder, S. Primates in Perspective. [S.l.]: Oxford University Press. pp. 107–120. ISBN 978-0-19-517133-4
- ↑ Rowe, N. (1996). The Pictorial Guide to the Living Primates. [S.l.]: Pogonias Press. p. 109. ISBN 978-0-9648825-0-8
- ↑ Melin, Amanda D.; Jack, Katherine M.; Fedigan, Linda; Mendez-Carvajal; Pedro G. (2016). Rowe, Noel; Myers, Marc, eds. All the World's Primates. [S.l.]: Pogonias Press. pp. 286–288. ISBN 9781940496061
- ↑ Morris, D.; Bruce, D. (2005). Primate Ethology. [S.l.]: Aldine Transaction. pp. 237–238. ISBN 978-0-202-30826-5
- ↑ Bezanson, L. (2006). «Ontogenetic Influences on Positional Behavior in Cebus and Alouatta». In: Estrada, A.; Garber, P.; Pavelka, M.; Luecke, L. New Perspectives in the Study of Mesoamerican Primates. New York: Springer. pp. 333–344. ISBN 978-0-387-25854-6
- ↑ a b c d Fragaszy, D.; Visalberghi, E.; Fedigan, L. (2004). «Life History and Demography». The Complete Capuchin. [S.l.]: Cambridge University Press. pp. 74–79
- ↑ Jack, K.; Fedigan, L. (2009). «Female dispersal in a female-philopatric species, Cebus capucinus.». Behaviour. 146 (4): 471–498. CiteSeerX 10.1.1.619.2612. doi:10.1163/156853909X404420
- ↑ a b c d e f g h i j k l m Perry, S.; Manson, J. (2008). Manipulative Monkeys: The Capuchins of Lomas Barbudal. Cambridge, MA: Harvard University Press. pp. 118, 145–154, 169–214, 229–241. ISBN 978-0-674-02664-3
- ↑ a b c Jack, K.; Fedigan, L. (2004). «Male dispersal patterns in white-faced capuchins, Cebus capucinus Part 1: patterns and causes of natal emigration». Animal Behaviour. 67 (4): 761–769. doi:10.1016/j.anbehav.2003.04.015
- ↑ a b Jack, K.; Fedigan, L. (2004). «Male dispersal patterns in white-faced capuchins, Cebus capucinus Part 2: Patterns and causes of secondary dispersal». Animal Behaviour. 67 (4): 771–782. doi:10.1016/j.anbehav.2003.06.015
- ↑ a b c Muniz, L.; Perry, S.; Manson, J.; Gilkenson, H.; Gros-Louis, J.; Vigilant, L. (2006). «Father-daughter inbreeding avoidance in a wild primate population.». Current Biology. 16 (5): 156–7. PMID 16527729. doi:10.1016/j.cub.2006.02.055
- ↑ a b c d e f g h Perry, S.; Manson, J.H.; Muniz, L.; Gros-Louis, J.; Vigilant, L. (2008). «Kin-biased Social Behaviour in Wild Adult Female White-faced Capuchins (Cebus capucinus)». Animal Behaviour. 76: 187–199. doi:10.1016/j.anbehav.2008.01.020
- ↑ a b Perry, S. (1996). «Female-female relationships in wild white-faced capuchin monkeys, Cebus capucinus.» (PDF). American Journal of Primatology. 40 (2): 167–182. doi:10.1002/(SICI)1098-2345(1996)40:2<167::AID-AJP4>3.0.CO;2-W. hdl:2027.42/38432
- ↑ Perry, S.; Manson, J.; Barrett, H.C. (2004). «White-faced capuchin monkeys exhibit triadic awareness in their choice of allies.». Animal Behaviour. 67: 165–170. doi:10.1016/j.anbehav.2003.04.005
- ↑ Bergstrom, M.; Fedigan, L.M. (2009). «Strength and stability of dominance hierarchies in female white-faced capuchins (Cebus capucinus) at Santa Rosa National Park, Costa Rica.». American Journal of Primatology. 71 (Suppl 1): 103
- ↑ a b Perry, S. (1997). «Male-female social relationships in wild white-faced capuchin monkeys, Cebus capucinus.». Behaviour. 134 (7): 477–510. doi:10.1163/156853997X00494
- ↑ a b c Perry, S. (1998). «Male-male social relationships in wild white-faced capuchins, Cebus capucinus.». Behaviour. 135 (2): 1–34. doi:10.1163/156853998793066384
- ↑ Manson, J.H.; Perry, S.; Parish, A.R. (1997). «Nonconceptive sexual behavior in bonobos and capuchins.». International Journal of Primatology. 18 (5): 767–786. doi:10.1023/A:1026395829818
- ↑ a b c d e Perry, S. (1996). «Intergroup encounters in wild white-faced capuchins, Cebus capucinus.». International Journal of Primatology. 17 (3): 309–330. doi:10.1007/BF02736624
- ↑ a b Gros-Louis, J.; Perry, S.; Manson, J.H. (2003). «Violent coalitionary attacks and intraspecific killing in wild capuchin monkeys (Cebus capucinus).». Primates. 44 (4): 341–346. PMID 12910384. doi:10.1007/s10329-003-0050-z
- ↑ Fedigan, L.; Jack, K. (2004). «The Demographic and Reproductive Context of Male Replacements in Cebus Capucinus» (PDF). Behaviour. 141 (6): 755–775. doi:10.1163/1568539042245178. Consultado em 14 de novembro de 2008
- ↑ a b c Jack, K.; Fedigan, L. (2006). «Dominance and Reproductive Success in Wild White-Faced Capuchins». In: Estrada, A.; Garber, P.; Pavelka, M.; Luecke, L. New Perspectives in the Study of Mesoamerican Primates. New York: Springer. pp. 367–382. ISBN 978-0-387-25854-6
- ↑ Jack; Fedigan (2006). «Why Be Alpha Male?». New Perspectives in the Study of Mesoamerican Primates. doi:10.1007/0-387-25872-8_18
- ↑ Manson JH, Gros-Louis J, Perry S (2004). «Three apparent cases of infanticide by males in wild white-faced capuchins (Cebus capucinus)». Folia Primatologica. 75 (2): 104–106. PMID 15010584. doi:10.1159/000076270
- ↑ a b Fedigan, L.M. (2003). «Impact of male takeovers on infant deaths, births, and conceptions in Cebus capucinus at Santa Rosa, Costa Rica.». International Journal of Primatology. 24 (4): 723–741. doi:10.1023/A:1024620620454
- ↑ Hrdy, S. (1974). «Male-male competition and infanticide among the langurs (Presbytis entellus) of Abu, Rajasthan.». Folia Primatologica. 22 (1): 19–58. PMID 4215710. doi:10.1159/000155616
- ↑ a b c d e f g h i j k l m n o Defler, T. (2004). Primates of Colombia. Bogotá, D.C., Colombia: Conservation International. pp. 227–235. ISBN 978-1-881173-83-0
- ↑ Fragaszy, D.; Visalberghi, E.; Fedigan, L. (2004). «Behavioral Ecology». The Complete Capuchin. [S.l.]: Cambridge University Press. pp. 38–39. ISBN 978-0-521-66768-5
- ↑ a b Crofoot, M.C.; Gilby, I.C.; Wikelski, M.C; Kays, RW (2008). «Interaction location outweighs the competitive advantage of numerical superiority in Cebus capucinus intergroup contests.». PNAS. 105 (2): 577–581. Bibcode:2008PNAS..105..577C. PMC 2206578. PMID 18184811. doi:10.1073/pnas.0707749105
- ↑ a b Rose, L.; Perry, S.; Panger, M.; Jack, K.; Manson, J.; Gros-Louis, J.; Mackinnin, K. (agosto de 2003). «Interspecific Interactions between Cebus capucinus and other Species: Data from Three Costa Rican Sites» (PDF). International Journal of Primatology. 24 (4): 780–785. doi:10.1023/A:1024624721363. Consultado em 4 de setembro de 2008. Arquivado do original (PDF) em 25 de fevereiro de 2009
- ↑ Boinski, S. (2000). «Social Manipulation Within and Between Troops Mediates Primate Group Movement». In: Boinski, S.; Garber, P. On the Move. [S.l.]: The University of Chicago Press. pp. 447–448. ISBN 978-0-226-06340-9
- ↑ Boinski, S. (abril de 1989). «Why don't Saimiri oerstedii and Cebus capucinus form mixed-species groups?». International Journal of Primatology (Submitted manuscript). 10 (2): 103–114. doi:10.1007/BF02736248
- ↑ Fragaszy, D.; Visalberghi, E.; Fedigan, L. (2004). «Community Ecology». The Complete Capuchin. [S.l.]: Cambridge University Press. p. 70. ISBN 978-0-521-66768-5
- ↑ Groves, C.P. (2005). Wilson, D. E.; Reeder, D. M, eds. Mammal Species of the World 3.ª ed. Baltimore: Johns Hopkins University Press. pp. 148–149. ISBN 978-0-8018-8221-0. OCLC 62265494
- ↑ «Capuchin monkeys caught abducting baby howlers: Cameras on a remote island in Panama captured the origin and spread of a bizarre "fad"». Max-Planck-Gesellschaft. Max Planck Institute. 19 de maio de 2025. Consultado em 1 de junho de 2025
- ↑ Goldsborough, Zoë; Crofoot, Margaret C.; Jacobson, Odd T.; Corewyn, Lisa; Rosario-Vargas, Eveyn del; León, Julián; Barrett, Brendan J. (19 de maio de 2025). «Rise and spread of a social tradition of interspecies abduction». Current Biology. 35 (10): 375–376. Consultado em 1 de junho de 2025
- ↑ Larson, Christina (19 de maio de 2025). «Caught on camera, capuchin monkeys kidnap howler monkey babies: Scientists have found evidence of monkey kidnappings in Panama». The Washington Post. Consultado em 1 de junho de 2025
- ↑ a b Chapman, C.; Fedigan, L. (1990). «Dietary Differences between Neighboring Cebus capucinus Groups: Local Traditions, Food Availability or Responses to Food Profitability?» (PDF). Folia Primatol. 54 (3–4): 177–186. PMID 2391047. doi:10.1159/000156442
- ↑ a b c d e Fragaszy, D.; Visalberghi, E.; Fedigan, L. (2004). «Behavioral Ecology». The Complete Capuchin. [S.l.]: Cambridge University Press. pp. 43–47. ISBN 978-0-521-66768-5
- ↑ Melin, Amanda D.; Fedigan, Linda Marie; Hiramatsu, Chihiro; Kawamura, Shoji (22 de setembro de 2007). «Polymorphic color vision in white-faced capuchins (Cebus capucinus): Is there foraging niche divergence among phenotypes?» (PDF). Behavioral Ecology and Sociobiology (em inglês). 62 (5): 663. ISSN 1432-0762. OCLC 437741616. doi:10.1007/s00265-007-0490-3. Consultado em 20 de julho de 2018
- ↑ MacKinnon, K. (2006). «Food Choice by Juvenile Capuchin Monkeys». In: Estrada, A.; Garber, P.; Pavelka, M.; Luecke, L. New Perspectives in the Study of Mesoamerican Primates. New York: Springer. pp. 354–360. ISBN 978-0-387-25854-6
- ↑ Luedtke, Karen (2010). «Ch 11». Costa Rica: Monkeys, Animal Behavior, Cognitive Neuroscience. [S.l.: s.n.] 109 páginas. ISBN 978-0-9832448-0-6
- ↑ a b David Attenborough (2003). Life of Mammals. BBC Video
- ↑ Perry S. Rose L. (1994). «Begging and transfer of coati meat by white-faced capuchin monkeys, Cebus capucinus» (PDF). Primates. 35 (4): 409–415. doi:10.1007/bf02381950. hdl:2027.42/41610
- ↑ Leake, D.; Dobson, R. (15 de abril de 2007). «Chimps Knocked Off Top of the IQ Tree». The Times. London. Consultado em 1 de setembro de 2008. Arquivado do original em 15 de maio de 2007
- ↑ Barrett, Brendan J.; Monteza-Moreno, Claudio M.; Dogandžić, Tamara; Zwyns, Nicolas; Ibáñez, Alicia; Crofoot, Margaret C. (2018). «Habitual stone-tool-aided extractive foraging in white-faced capuchins, Cebus capucinus». Royal Society Open Science. 5 (8). 181002 páginas. Bibcode:2018RSOS....581002B. PMC 6124021. PMID 30225086. doi:10.1098/rsos.181002
- ↑ Monteza-Moreno, Claudio M.; Dogandžić, Tamara; McLean, Kevin A.; Castillo-Caballero, Pedro L.; Mijango-Ramos, Zarluis; Del Rosario-Vargas, Evelyn; Crofoot, Margaret C.; Barrett, Brendan J. (1 de junho de 2020). «White-Faced Capuchin, Cebus capucinus imitator, Hammerstone and Anvil Tool Use in Riparian Habitats on Coiba Island, Panama». International Journal of Primatology (em inglês). 41 (3): 429–433. ISSN 1573-8604. doi:10.1007/s10764-020-00156-5
- ↑ Fragaszy, D.; Visalberghi, E.; Fedigan, L. (2004). «Capuchins Use Objects as Tools». The Complete Capuchin. [S.l.]: Cambridge University Press. pp. 173–183. ISBN 978-0-521-66768-5
- ↑ «Capuchin Monkey (Cebus capucinus)». The Rainforest Alliance. Consultado em 7 de fevereiro de 2009
- ↑ Blumenthal, D. (17 de junho de 1987). «Monkeys as Helpers To Quadriplegics At Home». The New York Times
- ↑ Pflum, M. (18 de março de 2000). «Earth Matters: Turkey struggles with national epidemic: primate smuggling». CNN. Consultado em 7 de fevereiro de 2009. Arquivado do original em 28 de setembro de 2006
- ↑ a b c d Henderson, C. (2000). Field Guide to the Wildlife of Costa Rica. [S.l.]: University of Texas Press. pp. 454–455. ISBN 978-0-292-73459-3
- ↑ a b Fragaszy, D.; Visalberghi, E.; Fedigan, L. (2004). «Social Interactions, Relationships and Social Structure». The Complete Capuchin. [S.l.]: Cambridge University Press. pp. 202–220. ISBN 978-0-521-66768-5
- ↑ Fragaszy, D.; Visalberghi, E.; Fedigan, L. (2004). «The Body». The Complete Capuchin. [S.l.]: Cambridge University Press. p. 102. ISBN 978-0-521-66768-5
- ↑ Carnegie, S.; Fedigan, L.; Ziegler, T. (2006). «Post-conceptive Mating in White-Faced Capuchins». In: Estrada, A.; Garber, P.; Pavelka, M.; Luecke, L. New Perspectives in the Study of Mesoamerican Primates. New York: Springer. pp. 387–405. ISBN 978-0-387-25854-6
- ↑ a b Di Fiore, A. (2009). «Genetic Approaches to the Study of Dispersal and Kinship in New World Primates». In: Garber, P.; Estrada, A.; Bicca-Marques, J.C.; Heymann, E.; Strier, K. South American Primates: Comparative Perspectives in the Study of Behavior, Ecology and Conservation. [S.l.]: Springer. pp. 222–223. ISBN 978-0-387-78704-6
- ↑ Panger, M.; Perry, S.; Rose, L.; Gros-Louis, J.; Vogel, E.; Mackinnon, C.; Baker, M. (2002). «Cross-Site Differences in Foraging Behavior of White-Faced Capuchins (Cebus capucinus)» (PDF). American Journal of Physical Anthropology. 119 (1): 52–66. PMID 12209573. doi:10.1002/ajpa.10103. Consultado em 6 de outubro de 2009. Arquivado do original (PDF) em 4 de junho de 2011
- ↑ Hunter, L.; Andrew, D. (2002). Watching Wildlife Central America. [S.l.]: Lonely Planet Publications. pp. 97, 100, 110, 130. ISBN 978-1-86450-034-9
- ↑ «Ometepe Island Info - Volcán Maderas». ometepeislandinfo.com (em inglês). Consultado em 5 de março de 2017
- ↑ DeGama, H.; Fedigan, L. (2006). «The Effects of Forest Fragment Age, Isolation, Size, Habitat Type, and Water Availability on Monkey Density in a Tropical Dry Forest». In: Estrada, A.; Garber, P.; Pavelka, M.; Luecke, L. New Perspectives in the Study of Mesoamerican Primates. New York: Springer. pp. 165–186. ISBN 978-0-387-25854-6
- ↑ Garber, P.; Estrada, A.; Pavelka, M. (2006). «Concluding Comments and Conservation Priorities». In: Estrada, A.; Garber, P.; Pavelka, M.; Luecke, L. New Perspectives in the Study of Mesoamerican Primates. New York: Springer. pp. 570–571. ISBN 978-0-387-25854-6
Leitura adicional
- Perry, S.; Manson, J. (2009). «11. Guapo: Innovation and Tradition in the Creation of Bond-Testing Rituals». Manipulative Monkeys: The Capuchins of Lomas Barbudal. Cambridge, MA and London, England: Harvard University Press. pp. 245–263
- Perry, S. (2011). «Social traditions and social learning in capuchin monkeys (Cebus)». Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 366 (1567): 988–996. PMC 3049088. PMID 21357221. doi:10.1098/rstb.2010.0317
- Perry, S.; Baker, M.; Fedigan, L.; Gros Louis, J.; Jack, K.; MacKinnon, Katherine C.; Manson, Joseph H.; Panger, M.; Pyle, K.; Rose, L. (2003). «Social Conventions in Wild White-faced Capuchin Monkeys: Evidence for Traditions in a Neotropical Primate». Current Anthropology. 44 (2): 241–268. doi:10.1086/345825
- Perry, S. E.; Barrett, B. J.; Godoy, I. (2017). «Older, sociable capuchins (Cebus capucinus) invent more social behaviors, but younger monkeys innovate more in other contexts». Proceedings of the National Academy of Sciences of the United States of America. 114 (30): 7806–7813. PMC 5544268. PMID 28739946. doi:10.1073/pnas.1620739114
- Perry, Susan (1995). Social relationships in wild white-faced capuchin monkeys, Cebus capucinus. Ann Arbor: University Microfilms International