Cardabiodon
Cardabiodon
| |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Cenomaniano-Turoniano, 95–91 Ma | |||||||||||||||||||
| Classificação científica | |||||||||||||||||||
| |||||||||||||||||||
| Espécie-tipo | |||||||||||||||||||
| †Cardabiodon ricki Siverson, 1999 | |||||||||||||||||||
| Espécies | |||||||||||||||||||
| |||||||||||||||||||
| Sinónimos | |||||||||||||||||||
| |||||||||||||||||||
![Dentes holótipos e parátipos de Cardabiodon venator do membro Fairport em Mosby [en], Montana.](./_assets_/0c70a452f799bfe840676ee341124611/Teeth_of_Cardabiodon_venator_from_the_Fairport_Member_of_the_Carlile_Shale_in_Mobsy%252C_Montana.png)
Cardabiodon ([ˌkɑːrdəbaɪəˈdɒn], significando "dente de Cardabia") é um gênero extinto de grandes tubarões da ordem Lamniformes que viveu há cerca de 95 a 91 milhões de anos, durante o Cenomaniano ao Turoniano do Cretáceo Superior.[2] Pertence à família Cardabiodontidae [en], única entre os tubarões da ordem Lamniformes devido a estruturas dentárias distintas, e inclui duas espécies: C. ricki e C. venator. Fósseis de Cardabiodon foram encontrados na Austrália, América do Norte, Inglaterra e Cazaquistão. Provavelmente era um tubarão com distribuição antitropical, habitando oceanos temperados neríticos e de mar aberto entre 40° e 60° de paleolatitude, semelhante ao moderno tubarão Lamna nasus.
Um dos maiores tubarões de sua época, Cardabiodon foi estimado em até 5,5 m de comprimento. Pode ter sido um superpredador em seu ecossistema, utilizando seus dentes grandes e robustos e sua capacidade de natação rápida para caçar uma variedade de animais marinhos, incluindo plesiossauros, ictiossauros e outros peixes grandes. Cardabiodon também possuía um corpo pesado e robusto. Cientistas conseguiram calcular parcialmente sua história de vida. Ao nascer, os indivíduos de Cardabiodon mediam entre 41 e 76 cm de comprimento e alcançavam a maturidade sexual entre cinco e sete anos de idade, mas não há estimativas conclusivas sobre sua longevidade máxima.
Descoberta e nomenclatura
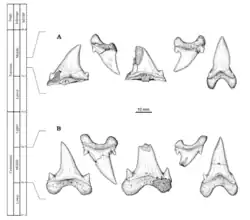
Cardabiodon foi descrito a partir de um fóssil associado descoberto na bacia de Carnarvon Sul, no siltito de Gearle, localizado em Cardabia [en], uma estação de gado [en] na Austrália Ocidental, pelo paleontólogo Mikael Siverson, que publicou suas descobertas em 1999.[2] Essa localidade é datada do Cenomaniano do Cretáceo Superior, cerca de 96 a 94,7 milhões de anos atrás.[3] O espécime, catalogado como WAM 96.4.45, consistia originalmente em uma associação de cem dentes e quinze vértebras, representando um tubarão subadulto; expedições posteriores ao mesmo local recuperaram mais quatro dentes pertencentes ao mesmo espécime.[4] Siverson reconheceu o espécime como holótipo de um novo gênero e atribuiu a ele o nome científico Cardabiodon ricki. O nome genérico Cardabiodon combina o nome do local Cardabia com o grego antigo ὀδών (odṓn, "dente"), significando "dente de Cardabia".[2] O epíteto específico ricki significa "de Rick", em homenagem a Rick French, proprietário da estação Cardabia.[2] Em 2005, a segunda espécie, C. venator, foi descrita a partir de espécimes-tipo consistindo em 37 dentes recuperados de uma localidade do membro Fairport, próximo a Mosby [en], Montana, uma formação datada de cerca de 92 a 91 milhões de anos atrás. O epíteto específico venator é uma palavra em latim que significa "caçador", referindo-se à presumida posição de C. venator como superpredador ou próximo ao topo da cadeia alimentar.[3]
Antes da descrição de Siverson para Cardabiodon, seus dentes foram descritos no passado sob dois táxons agora sinonimizados. A descrição original foi feita em 1957 pelo paleontólogo soviético Leonid Glickman, que descreveu o táxon Pseudoisurus tomosus com base em quatro dentes do oblast de Saratov.[5] Anteriormente, em 1894, Sir Arthur Smith Woodward descreveu uma associação de 25 dentes de giz cretáceo próximo a Maidstone, Inglaterra, sob o táxon Lamna appendiculata (Cretalamna appendiculata),[6] que, em 1977, o paleontólogo belga Jaques Herman concluiu pertencer a uma nova espécie, criando o táxon Cretolamna woodwardi.[7] Em 1996, Siverson sinonimizou Cretolamna woodwardi com Pseudoisurus tomosus após constatar que seus espécimes-tipo referidos pertenciam à mesma espécie.[8] Mais tarde, em 1999, Siverson reavaliou a taxonomia de Pseudoisurus tomosus e concluiu que seus síntipos estavam perdidos, tornando-o um nomen dubium. Ele também descobriu que o antigo Cretolamna woodwardi representava dois gêneros distintos de tubarões, o que provavelmente também ocorria com Pseudoisurus tomosus. O primeiro foi C. ricki (que ele já havia descrito usando WAM.96.4.45), e o outro foi atribuído ao táxon Dwardius woodwardi.[2]
Descrição
Anatomia geral

C. ricki foi estimado em até 5,5 m de comprimento por meio de comparações vertebrais com um tubarão-branco moderno do mesmo tamanho, enquanto C. venator foi estimado em até 5 m com base em análises dentárias,[4] tornando Cardabiodon um dos maiores tubarões conhecidos.[2] O registro fóssil é escasso, consistindo atualmente de dentes, vértebras e escamas, o que é comum, já que a cartilagem dos tubarões não se preserva bem durante a fossilização, embora as vértebras possam ser preservadas se endurecidas por calcificação.[9]
A pele de Cardabiodon era coberta por escamas revestidas com 6 a 8 sulcos paralelos, cada um com quilhas. Em um indivíduo medindo de 2,5 a 3,8 m de comprimento, essas escamas variavam de 0,3 a 0,7 mm em suas dimensões máximas. Escamas com sulcos e quilhas são eficientes na redução do arrasto hidrodinâmico e estão correlacionadas com a capacidade de natação do tubarão que as possui, sendo comuns em tubarões nadadores rápidos, como os lamnídeos. Isso sugere que Cardabiodon também era capaz de natação rápida e, portanto, de perseguir presas ágeis.[9]
As vértebras eram robustas. As duas camadas mais externas de uma vértebra de um tubarão da ordem Lamniformes e, associadas à relação entre comprimento e peso, eram mais espessas que as de outros grandes tubarões predadores da ordem Lamniformes, sugerindo que Cardabiodon era significativamente mais pesado e robusto, embora ainda não tenha sido feita uma estimativa de seu peso. A coluna vertebral era rígida e em forma de fuso, o que seria pouco flexível, mas eficiente para natação rápida.[4] Cardabiodon também tinha um corpo maior em relação aos seus dentes.[9]
Dentição
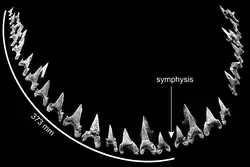
Cardabiodon possuía as características dentárias básicas de um tubarão da ordem Lamniformes.[4] Sua estrutura dentária era heterodonte, apresentando várias morfologias de dentes.[2] Características diagnósticas dos dentes de Cardabiodon incluem raízes fortemente bilabiais, coroas robustas que são quase simetricamente erguidas ou curvadas distalmente, pescoços dentários visíveis e grandes, bordas de corte não serrilhadas e cúspides laterais.[2][3][9][10] O lado lingual, voltado para a boca, apresenta uma raiz massivamente convexa com pequenos poros nutritivos.[10] A coroa no lado labial, voltada para fora, exibe protuberância.[1]
Os dentes de Cardabiodon são de tamanho moderado.[11] Os maiores exemplos de dentes de C. ricki provêm do holótipo.[a] Seus dentes anteriores medem até 3,67 cm na altura inclinada máxima, enquanto os primeiros dentes lateroposteriores inferiores são maiores, medindo até 3,98 cm na altura inclinada máxima. Os dentes de C. venator são ligeiramente menores, com o maior dente conhecido sendo um anterior medindo 3,26 cm na altura inclinada máxima, mas são muito mais volumosos e espessos. C. venator também possui cúspides laterais significativamente mais curtas em comparação com C. ricki.[3]
Siverson reconstruiu uma fórmula dentária artificial com base nos 104 dentes do holótipo de C. ricki,[2][4] que ele revisou em um artigo de 2015 após concluir que superestimou o número de dentes ausentes na mandíbula inferior.[4] A fórmula dentária baseada nesses estudos é .[b][4][9] Isso significa que Cardabiodon possuía principalmente quatro tipos de dentes em sua dentição, ordenados da frente para trás: sinfisiais, anteriores, intermediários e lateroposteriores. A estrutura dentária de Cardabiodon é única e incomparável com qualquer outro tubarão extinto ou existente conhecido. Na mandíbula inferior, o tamanho dos dentes aumenta abruptamente e atinge o pico no primeiro dente lateroposterior. Isso é contrário às estruturas dentárias mais genéricas de tubarões da ordem Lamniformes, onde o tamanho dos dentes diminui gradualmente da região anterior para a posterior (com exceção dos dentes sinfisiais e intermediários menores).[4] Os dentes inferiores também são mais largos que os dentes da mandíbula superior, enquanto nos tubarões genéricos da ordem Lamniformes os dentes superiores são mais largos.[9] A circunferência de mordida da mandíbula inferior do holótipo de C. ricki foi medida em 746 mm.[4]

Uma fórmula dentária alternativa foi reconstruída a partir de um conjunto de dentes desarticulados identificado como Cardabiodon sp. do xisto de Carlile no Kansas, que é . O estudo que criou essa fórmula buscou ser mais consistente com a estrutura genérica de tubarões da ordem Lamniformes, diferente da reconstruída por Siverson, que os cientistas expressaram com cautela devido a inconsistências com a estrutura dentária genérica de tubarões da ordem Lamniformes. No entanto, ambas são hipotéticas devido à natureza desarticulada dos dentes fósseis usados.[9]

Classificação e evolução

Cardabiodon era um tubarão único devido a sua estrutura dentária distinta. Ainda assim, é classificado na ordem Lamniformes devido à posse de características vertebrais e dentárias básicas exclusivas de tubarões desta ordem. Para refletir a estrutura dentária única de Cardabiodon, Siverson criou a família Cardabiodontidae e colocou o táxon, juntamente com um tubarão extinto do Cenozoico, Parotodus [en], nela.[2] No entanto, ele posteriormente expressou uma diminuição na justificativa para incluir Parotodus na família, citando principalmente um intervalo fóssil de cerca de 60 milhões de anos entre ele e Cardabiodon.[3] Outro tubarão do Cretáceo Superior, Dwardius [en], cujos dentes são muito semelhantes aos de Cardabiodon, também foi colocado na família. Foi proposta uma relação próxima entre os dois,[13] e a possibilidade de serem gêneros sinônimos foi levantada por um estudo de 2010 liderado pelo paleontólogo Todd D. Cook. No entanto, Siverson considerou improvável essa proposta, notando tendências evolutivas contraditórias, como a diminuição gradual de cúspides laterais em Cardabiodon ao longo do tempo, em comparação com o aumento gradual em Dwardius, mas concordou que os dois gêneros eram estreitamente relacionados.[11] O posicionamento filogenético da família Cardabiodontidae é incerto e ainda não foi estabelecido.[3] Um dente de C. ricki recuperado de depósitos em Alberta, datado de cerca de 93 milhões de anos, foi descoberto contendo características geralmente encontradas em C. venator, sugerindo uma relação cronospecífica entre as duas espécies, com o dente representando um morfotipo transicional.[1][4]
Embora a família Cardabiodontidae contenha apenas os dois gêneros Cardabiodon e Dwardius,[13] vários fósseis de tubarões não descritos de períodos anteriores são conhecidos. O mais notável inclui um conjunto associado de dentes e vértebras muito grandes datadas do Albiano, cerca de 105 milhões de anos, da formação Toolebuc [en] em Queensland, pertencente a um indivíduo estimado em 8 a 9 m de comprimento.[14] Em uma discussão de blog, Siverson observou que os dentes desse espécime do Albiano são menos robustos que os de outros tubarões predadores do Cretáceo, como o Cretoxyrhina. Ele sugeriu uma tendência evolutiva da família Cardabiodontidae marcada por uma redução no tamanho e comprimento do corpo, uma redução no número de dentes por linha maxilar, um aumento na robustez dos dentes (incluindo maior robustez da raiz e alargamento da coroa) e uma redução das cúspides laterais.[12]
Paleobiologia
História de vida

Como todos os tubarões da ordem Lamniformes, Cardabiodon desenvolvia um anel de crescimento em suas vértebras a cada ano, e sua idade é determinada pela contagem desses anéis. No entanto, poucos indivíduos de Cardabiodon foram datados devido à raridade de vértebras bem preservadas. A ontogenia de C. ricki foi calculada usando as vértebras de três indivíduos pré-adultos, além de um dente isolado de um recém-nascido.[4] Quando comparado com os registros ontogenéticos de Cretoxyrhina mantelli e Archaeolamna kopingensis, as vértebras de C. ricki sugeriram que o comprimento ao nascer era menor que o dos dois tubarões, variando entre 41 e 76 cm. Ele crescia a uma taxa média de 29,9 cm por ano, em comparação com a taxa de crescimento média mais rápida de C. mantelli (40,7 cm por ano) e a mais lenta de A. kopingensis (6,2 cm por ano).[15] Todas as três vértebras mostraram treze anéis de crescimento, indicando que os indivíduos viveram cerca de 13 anos. No entanto, uma taxa de crescimento mais lenta que a de C. mantelli sugere uma longevidade máxima indeterminável superior a 13 anos, e que as idades encontradas nos espécimes resultaram de morte prematura. Por volta do quinto ao sétimo anel de crescimento nas três vértebras, há uma mudança no crescimento, sugerindo que C. ricki alcançava a maturidade sexual entre 5 e 7 anos de idade.[4]
Paleoecologia
Distribuição antitropical
Fósseis de Cardabiodon foram encontrados no Canadá, no centro-oeste dos Estados Unidos, na Inglaterra, na Austrália Ocidental[4] e no Cazaquistão.[2] No passado, foi sugerido que Cardabiodon poderia ter uma distribuição antitropical, já que essas localidades estavam entre 40° e 60° de paleolatitude, semelhante à faixa de Lamna nasus, que existe apenas entre 30° e 60° de latitude. Isso foi confirmado por um estudo de 2010 que encontrou que as faixas latitudinais e de temperatura da superfície do mar entre os dois eram quase idênticas. O estudo também descobriu que os fósseis de maior latitude de Cardabiodon foram datados pouco antes do período mais quente do Cretáceo Superior, cerca de 93 milhões de anos, conhecido como Máximo Térmico Cenomaniano-Turoniano. Isso sugere um deslocamento de distribuição para o norte devido ao aumento das temperaturas e ambientes tropicais. Determinou-se que a faixa de temperatura da superfície do mar de Cardabiodon era de 17,5 a 24,2 °C.[1] Propôs-se que Cardabiodon poderia se mover de um hemisfério para o outro cruzando águas mais profundas e, portanto, mais frias. Isso foi observado em tubarões-peregrinos modernos, outro tubarão antitropical, que migra entre hemisférios seguindo caminhos isotérmicos mais frios em águas tropicais profundas, onde as temperaturas são semelhantes à sua faixa normal. Embora nenhum fóssil de Cardabiodon tenha sido encontrado fora de sua faixa inferida, essa proposta é apoiada pela ocorrência de dentes de Cardabiodon em depósitos de águas profundas em torno de sua menor paleolatitude.[1]
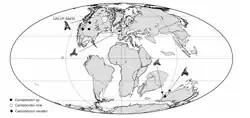
Habitat
A distribuição antitropical de Cardabiodon sugere que ele vivia principalmente em águas temperadas e de mar aberto, mas moderadamente profundas, do interior à zona nerítica média. A maioria de seus fósseis é proveniente da América do Norte.[4] Na América do Norte, os fósseis de C. venator são encontrados principalmente na metade sul do Mar Interior Ocidental, que tinha um clima temperado mais quente; eles são raramente encontrados na metade norte.[16] Embora tenha vivido em temperaturas marítimas mais frias de 17,5 a 24,2 °C, Cardabiodon foi contemporâneo ao Máximo Térmico Cenomaniano-Turoniano,[1] que levou a uma mudança na biodiversidade e ao surgimento e radiação de uma nova fauna, como os mosassauros. Localidades do Cenomaniano no Mar Interior Ocidental renderam vários vertebrados marinhos que coexistiram com Cardabiodon, que o tubarão, presumivelmente como superpredador, pode ter caçado. Estes incluem muitos tubarões, como os da ordem Lamniformes Cretodus [en], Cretalamna, Protolamna [en] e Cretoxyrhina; membros da família Anacoracidae como Squalicorax; e hibodontes como Ptychodus [en] e Hybodus; grandes peixes ósseos como Protosphyraena, Pachyrhizodus [en], Enchodus [en] e Xiphactinus; aves marinhas como Pasquiaornis [en] e Ichthyornis; répteis marinhos como elasmossaurídeos e policotilídeos plesiossauros; o pliossauro Brachauchenius lucasi [en], tartarugas marinhas protostegídeas e dolicossaurídeos como Coniasaurus crassidens [en].[16] O siltito de Gearle, na Austrália Ocidental, era dominado principalmente por Cretalamna, mas outros tubarões, como Squalicorax, Archaeolamna, Paraisurus [en], Notorhynchus, Leptostyrax [en] e Carcharias, estavam presentes.[17] Restos fragmentados de pliossauros também são conhecidos da formação. Outras partes da Austrália Ocidental também eram habitadas durante o Cenomaniano por ictiossauros como Platypterygius [en].[18] Faunas bentônicas eram raras nessa época, especialmente nas águas mais ao norte habitadas por Cardabiodon, devido principalmente aos baixos níveis de oxigênio[16] causados pelo Evento Anóxico Cenomaniano-Turoniano, que levou à extinção de até 27% de todos os invertebrados marinhos.[19]
Áreas de berçário
Como muitos tubarões modernos, Cardabiodon utilizava áreas de berçário para dar à luz e criar seus filhotes, idealmente em águas rasas que oferecem proteção contra predadores naturais. Uma área do xisto de Carlile, próximo a Mosby, Montana, foi identificada como um habitat de berçário devido à alta prevalência de fósseis de filhotes de Cardabiodon.[3] Outras localidades na região do Mar Interior Ocidental da América do Norte, incluindo a formação Kaskapau no noroeste de Alberta e o calcário Greenhorn no centro de Kansas, também relataram fósseis de filhotes.[20] Como tais fósseis são amplamente ausentes em outros lugares, isso levou à sugestão de que o Mar Interior Ocidental como um todo pode ter servido como uma área de berçário para Cardabiodon.[4]
Notas
- ↑ Em um post de blog, Siverson afirmou que os maiores dentes de Cardabiodon encontrados eram até 15% maiores que os do holótipo.[12] No entanto, essas descobertas não foram publicadas validamente.
- ↑ Usando a interpretação de Dickerson et al. (2012) da fórmula, que utiliza uma terminologia mais comumente usada.[9]
Referências
- ↑ a b c d e f Todd D. Cook; Mark V. H. Wilson; Michael G. Newbrey (2010). «The first record of the large Cretaceous lamniform shark Cardabiodon ricki from North America and a new empirical test for its presumed antitropical distribution». Journal of Vertebrate Paleontology. 30 (3): 643–649. JSTOR 40666186. doi:10.1080/02724631003758052
- ↑ a b c d e f g h i j k Mikael Siverson (1999). «A new large lamniform shark from the uppermost Gearle Siltstone (Cenomanian, Late Cretaceous) of Western Australia». Transactions of the Royal Society of Edinburgh: Earth Sciences. 90 (1): 49–66. doi:10.1017/S0263593300002509
- ↑ a b c d e f g Mikael Siverson; Johan Lindgren (2005). «Late Cretaceous sharks Cretoxyrhina and Cardabiodon from Montana, USA» (PDF). Acta Palaeontologica Polonica. 50 (2): 301–314
- ↑ a b c d e f g h i j k l m n o Michael G. Newbrey; Mikael Siverson; Todd D. Cook; Allison M. Fotheringham; Rebecca L. Sanchez (2015). «Vertebral Morphology, Dentition, Age, Growth, and Ecology of the Large Lamniform Shark Cardabiodon ricki». Acta Palaeontologica Polonica. 60 (4): 877–897. doi:10.4202/app.2012.0047

- ↑ Leonid S. Glickman (1957). «О генетической связи семейства Lamidae и Odontaspidae и новых родах верхнемеловых ламнид» [Genetic relations of the Lamnidae and Odontaspidae and new genera of lamnids from the Upper Cretaceous] (PDF). Trudy Geologicheskogo Muzeia Akademiia Nauk SSSR (em russo). 1: 110-117
- ↑ Arthur S. Wooward (1894). «Notes on the sharks' teeth from British Cretaceous formations». Proceedings of the Geologists' Association. 13 (6): 190–200. doi:10.1016/S0016-7878(94)80009-4
- ↑ Jaques Herman (1977). Les Sélaciens des terrains néocrétacés & paléocènes de Belgique & des contrées limitrophes : éléments d'une biostratigraphie intercontinentale (PDF) (em francês). Bruxelas: Geological Survey of Belgium. pp. 207–209. OCLC 612294375
- ↑ Mikael Siverson (1996). «Lamniform sharks of the mid Cretaceous Alinga Formation and Beedagong Claystone, Western Australia» (PDF). Palaeontology. 39 (4): 813-849
- ↑ a b c d e f g h Ashley A. Dickerson; Kenshu Shimada; Brian Reilly; Cynthia R. Rigsby (2012). «New data on the Late Cretaceous cardabiodontid lamniform shark based on an associated specimen from Kansas». Transactions of the Kansas Academy of Science. 115 (3 & 4): 125–134. doi:10.1660/062.115.0305
- ↑ a b Alexander D. Meglei; Kenshu Shimada; James I. Kirkland (2013). «Fossil Vertebrates from the Middle Graneros Shale (Upper Cretaceous: Middle Cenomanian) in Southeastern Nebraska». Transactions of the Kansas Academy of Science. 116 (3–4): 129–136. doi:10.1660/062.116.0304
- ↑ a b Mikael Siverson; David J. Ward; Johan Lindgren; L. Scott Kelley (2012). «Mid-Cretaceous Cretoxyrhina (Elasmobranchii) from Mangyshlak, Kazakhstan and Texas, USA». Alcheringa. 37 (1): 87–104. doi:10.1080/03115518.2012.709440
- ↑ a b Mikael Siverson (22 de novembro de 2018). «Cardabiodon size, HELP!». The Fossil Forum. Consultado em 4 de março de 2019
- ↑ a b Mikael Siverson; Marcin Machalski (2017). «Late late Albian (Early Cretaceous) shark teeth from Annopol, Poland». Alcheringa: An Australasian Journal of Palaeontology. 41 (4): 433–463. doi:10.1080/03115518.2017.1282981
- ↑ Mikael Siverson (2012). Lamniform Sharks: 110 Million Years of Ocean Supremacy. Royal Tyrrell Museum of Palaeontology. Cópia arquivada em 15 de dezembro de 2021
- ↑ Joseph A. Fredrickson; Joshua E. Cohen; Jeff L. Berry (2016). «Ontogeny and life history of a large lamniform shark from the Early Cretaceous of North America». Cretaceous Research. 59 (1): 272–277. doi:10.1016/j.cretres.2015.11.007
- ↑ a b c Stephen L. Cumbaa; Kenshu Shimada; Todd D. Cook (2010). «Mid-Cenomanian vertebrate faunas of the Western Interior Seaway of North America and their evolutionary, paleobiogeographical, and paleoecological implications». Palaeogeography, Palaeoclimatology, Palaeoecology. 295 (1–2): 199–214. Bibcode:2010PPP...295..199C. doi:10.1016/j.palaeo.2010.05.038
- ↑ Mikael Siverson (1997). «Sharks from the mid-Cretaceous Gearle Siltstone, Southern Carnarvon Basin, Western Australia». Journal of Vertebrate Paleontology. 17 (3): 453–465. JSTOR 4523829. doi:10.1080/02724634.1997.10010995
- ↑ Benjamin Kear (2003). «Cretaceous marine reptiles of Australia: a review of taxonomy and distribution» (PDF). Cretaceous Research. 24 (2003): 277–303. doi:10.1016/S0195-6671(03)00046-6
- ↑ In Brief (16 de junho de 2008). «Submarine eruption bled Earth's oceans of oxygen». New Scientist. Consultado em 24 de novembro de 2018. Cópia arquivada em 6 de janeiro de 2014
- ↑ Todd D. Cook; Mark V.H. Wilson; Alison M. Murray; A. Guy Plint; Michael G. Newbrey; Michael J. Everhart (2013). «A high latitude euselachian assemblage from the early Turonian of Alberta, Canada». Journal of Systematic Palaeontology. 11 (5): 555–587. doi:10.1080/14772019.2012.707990