Agathaeromys
Agathaeromys
| |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Pleistoceno Médio-Superior ([[Ensenadano [en]]]-Lujaniano) ~0,9–0,23 Ma | |||||||||||||||||||
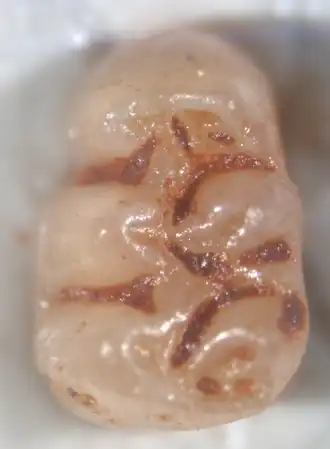 Primeiro molar superior esquerdo de Agathaeromys donovani | |||||||||||||||||||
| Classificação científica | |||||||||||||||||||
| |||||||||||||||||||
| Espécie-tipo | |||||||||||||||||||
| Agathaeromys donovani Zijlstra, Madern, e Van den Hoek Ostende, 2010 | |||||||||||||||||||
| Distribuição geográfica | |||||||||||||||||||
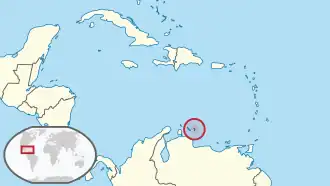 Localização da ilha de Bonaire, o único lugar onde foram encontrados fósseis de Agathaeromys.
| |||||||||||||||||||
| Espécies | |||||||||||||||||||
| |||||||||||||||||||
Agathaeromys[1] é um gênero extinto de roedores da tribo Oryzomyini do Pleistoceno de Bonaire, Antilhas Neerlandesas. Duas espécies são conhecidas, que diferem em tamanho e em alguns detalhes da morfologia dentária. A maior, A. donovani, a espécie-tipo, é conhecida por centenas de dentes, provavelmente com 900.000 a 540.000 anos de idade, encontrados em quatro localidades. A menor, A. praeuniversitatis, é conhecida por 35 dentes encontrados em um único sítio fóssil, com provável idade entre 540.000 e 230.000 anos.
Embora o material de Agathaeromys tenha sido descrito pela primeira vez em 1959, o gênero só foi formalmente nomeado e diagnosticado em 2010. Provavelmente pertence ao "clado D" dentro do grupo Oryzomyini, junto com várias outras espécies insulares. Os molares de ambas as espécies possuem várias cristas acessórias além das principais cúspides. Além de algumas diferenças nas características da superfície de mastigação dos molares, A. donovani apresenta mais raízes em seus molares inferiores do que A. praeuniversitatis.
Etimologia
O nome Agathaeromys combina as palavras gregas ἀγαθός agathos ("bom"), ἀήρ aêr ("ar") e μυς mys ("rato"), em referência ao nome da ilha de Bonaire e ao "ar fresco" que as contribuições de Marcelo Weksler e colegas trouxeram à classificação dos Oryzomyini.[1] O nome donovani homenageia Stephen Donovan [en] por suas contribuições ao conhecimento científico do Caribe,[2] e praeuniversitatis refere-se ao Pre-University College da Universidade de Leiden, que proporcionou a Jelle Zijlstra a oportunidade de participar do projeto que levou à identificação de Agathaeromys.[3]
Taxonomia
O material de Agathaeromys foi descrito pela primeira vez por Dirk Albert Hooijer [en] em 1959, no mesmo artigo que nomeou o rato gigante extinto de Curaçau, Megalomys curazensis [en]. Hooijer descreveu alguns dentes e mandíbulas fósseis de Fontein, Bonaire, como uma espécie indeterminada de Thomasomys ("Thomasomys sp.") e os considerou semelhantes a espécies agora classificadas como Delomys.[4] Em sua monografia de 1974 sobre a geologia das Ilhas ABC (Aruba, Bonaire e Curaçau — três ilhas neerlandesas ao noroeste da Venezuela), Paul Henri de Buisonjé listou Thomasomys sp. de outros sítios fósseis em Bonaire e mencionou adicionalmente Oryzomys sp. de um sítio diferente em Bonaire, Seroe Grandi.[5] Embora o material de Bonaire representasse um dos poucos registros fósseis de Thomasomys, ele foi raramente mencionado na literatura.[6]
Em 2010, Jelle Zijlstra, Anneke Madern e Lars van den Hoek Ostende revisaram o material. Eles consideraram improvável que "Thomasomys" de Bonaire pertencesse ao gênero sul-brasileiro Delomys ou a Thomasomys, que ocorre apenas nas montanhas dos Andes.[6] Por meio de uma análise cladística da subfamília Sigmodontinae, forneceram evidências de que o material de Bonaire pertencia à tribo Oryzomyini, e não a Thomasomyini (que inclui Thomasomys).[7] Realizaram outra análise cladística focada em Oryzomyini, que sugeriu que "Thomasomys" de Bonaire e o material de Seroe Grandi (Oryzomys sp. de De Buisonjé) eram intimamente relacionados, mas distintos de qualquer gênero de Oryzomyini conhecido.[8] Assim, nomearam um novo gênero de Oryzomyini, Agathaeromys, com duas espécies: Agathaeromys donovani (espécie-tipo) para o material anteriormente identificado como Thomasomys; e Agathaeromys praeuniversitatis para o material de Seroe Grandi.[9]
Embora Zijlstra e colegas não tenham determinado precisamente a posição de Agathaeromys dentro de Oryzomyini, seus resultados sugerem que ele está próximo à base do "clado D", um dos principais subgrupos de Oryzomyini. Este clado contém várias espécies exclusivas de ilhas, incluindo membros de Aegialomys, Agathaeromys, Megalomys, Nesoryzomys, Noronhomys, Oryzomys e Pennatomys. Zijlstra e colegas sugeriram que isso está relacionado à alta proporção de espécies semiaquáticas e não florestais no clado D — a maioria dos outros membros de Oryzomyini habita florestas.[10] No entanto, estudos filogenéticos subsequentes baseados em variações do mesmo conjunto de dados usado por Zijlstra e colegas não corroboraram essa colocação. Na descrição de Megalomys georginae [en] em 2012, Turvey e colegas posicionaram Agathaeromys fora dos principais grupos de Oryzomyini.[11] Zijlstra colocou Agathaeromys dentro do "clado C" em um clado com Oligoryzomys victus e uma espécie fóssil não descrita de Oligoryzomys de Aruba em seu artigo de 2012 nomeando Dushimys.[12]
No geral, Oryzomyini inclui mais de cem espécies em cerca de trinta gêneros.[13] Oryzomyini é uma das várias tribos dentro da subfamília Sigmodontinae da família Cricetidae, que abrange centenas de outras espécies de roedores, principalmente pequenos, distribuídos sobretudo na Eurásia e nas Américas.[14]
Descrição
Agathaeromys é caracterizado principalmente com base nas características dos dentes molares. Como é comum em roedores da superfamília Muroidea, há três molares em ambos os lados da maxila superior (referidos como M1, M2 e M3, da frente para trás) e da mandíbula inferior (referidos como m1, m2 e m3).[6] Agathaeromys é geralmente semelhante a outros membros de Oryzomyini, mas difere de outros gêneros em vários detalhes das coroas molares.[15] Agathaeromys donovani (comprimento do M1: 2,03–2,84 mm) é substancialmente maior que A. praeuniversitatis (comprimento do M1: 1,77–1,94 mm).[16]
Dentição superior
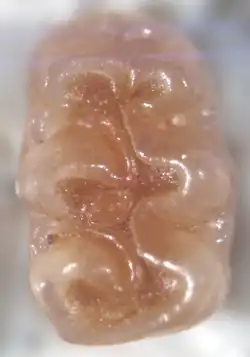
A cúspide frontal do M1, o anterocone, é dividida em duas cúspides menores por uma indentação. Em A. donovani, uma crista geralmente conecta as duas cúspides na margem frontal do dente, de modo que a indentação que as separa é fechada na frente (um fosseta anteromediana), mas em A. praeuniversitatis, ela é aberta para a frente (um flexo anteromediano). Uma crista adicional, o anterolofo, está presente atrás do anterocone. Mais atrás, há um par de cúspides — o protocone no lado lingual (interno) e o paracone no lado labial (externo). Uma crista parte do paracone e se conecta à parte frontal ou média do protocone. Atrás do paracone, a crista chamada mesolofo está presente; uma crista adicional geralmente conecta os dois. Na parte posterior do dente, há duas cúspides grandes adicionais — o hipocone (lingual) e o metacone (labial) — e uma crista proeminente, o posterolofo, parte do hipocone e está localizada atrás do metacone. Em A. donovani, o metacone geralmente está conectado ao posterolofo, mas em A. praeuniversitatis, ele está diretamente conectado ao hipocone. Os vales que separam as cúspides se estendem das margens lingual e labial até aproximadamente a linha média do molar. Além de uma grande raiz na frente e duas grandes raízes atrás (uma labial, uma lingual), há uma pequena raiz labial adicional.[17]
Não há anterocone no M2, mas um grande anterolofo está presente à frente do paracone. Há um protoflexo — uma indentação na coroa à frente do protocone. Como no M1, o paracone está conectado à parte frontal ou média do protocone, o mesolofo está bem desenvolvido, e os vales se encontram na linha média do dente. Há três raízes: duas no lado labial e uma no lado lingual.[17]
O M3 é um dente pequeno e triangular. Há uma grande bacia no centro, e um mesolofo está presente. Na parte posterior do dente, há um posterolofo distinto. Em A. donovani, há duas raízes na frente (labial e lingual) e uma atrás. Não há M3 de A. praeuniversitatis com raízes preservadas.[17]
Dentição inferior
O anteroconídeo — a cúspide frontal do m1 — é geralmente dividido em dois por uma indentação central em A. donovani, mas essa indentação está ausente em A. praeuniversitatis. Atrás do anteroconídeo, está o par de cúspides protoconídeo–metaconídeo. Há um cíngulo anterolabial — uma crista na margem labial frontal, à frente do protoconídeo. Há uma crista longa atrás do metaconídeo, um mesolofídeo. Geralmente, não há uma crista correspondente (um ectolofídeo) atrás do protoconídeo. Outro par de cúspides — o hipoconídeo e o entoconídeo — está localizado na parte posterior do dente. O entoconídeo, a cúspide lingual do par, está orientado para a frente. Há sempre uma grande raiz na frente do dente e outra atrás. A. donovani geralmente tem uma pequena raiz labial entre as duas raízes grandes e, frequentemente, outra pequena raiz no lado lingual, mas apenas um dos quatro m1s de A. praeuniversitatis possui a pequena raiz labial.[17]
Não há anteroconídeo no m2, e o dente não possui uma crista adicional (o anterolofídeo) à frente do metaconídeo, mas há um cíngulo anterolabial à frente do protoconídeo. Há um mesolofídeo. Além de uma grande raiz atrás, há duas raízes na frente em A. donovani, que às vezes estão parcialmente fundidas, mas apenas uma em A. praeuniversitatis.[18]
O cíngulo anterolabial e o anteroconídeo estão ausentes no m3. O mesolofídeo geralmente está ausente. Como no m2, há duas raízes na frente em A. donovani e apenas uma em A. praeuniversitatis, mas as raízes frontais geralmente estão fundidas em A. donovani.[19]
Mandíbulas

A maxila (mandíbula superior) é conhecida apenas para A. donovani. Nestes fósseis, a margem posterior do forame incisivo (uma abertura no palato) está aproximadamente no mesmo nível que a frente do M1, e a margem posterior da placa zigomática (uma placa óssea na lateral do crânio, conectada ao arco zigomático) também está próxima à frente do M1.[3] Mandíbulas (maxilares inferiores) de ambas as espécies são conhecidas. O forame mentual (uma abertura na frente do osso da mandíbula) abre-se para o lado labial do osso, exceto em uma mandíbula de A. praeuniversitatis, na qual a abertura está localizada mais acima. Há um processo capsular bem desenvolvido — uma elevação no osso que abriga a raiz do incisivo inferior. As cristas masseterianas (duas cristas no lado labial do osso que ancoram alguns dos músculos mastigatórios) estão unidas em uma única crista na direção da frente e alcançam um ponto abaixo da frente do m1.[20]
Uma única mandíbula de Porto Spanjo (um dos sítios onde fósseis de A. donovani foram encontrados), sem molares preservados, difere de todas as mandíbulas de Agathaeromys e é considerada como representando um roedor da subfamília Sigmodontinae diferente e desconhecido. Essa mandíbula é mais delgada que as de A. donovani, tem um diastema mais curto entre os incisivos e os molares, possui o incisivo menos deslocado lingualmente em relação aos molares e tem mais raízes sob os molares, conforme indicado pelos alvéolos dentários preservados.[21]
Idade e distribuição
Agathaeromys donovani é conhecido de quatro sítios fósseis — Fontein, Porto Spanjo, Barcadera-Karpata e "80 m acima do nível do mar" — que forneceram, respectivamente, 259, 148, 54 e 5 molares.[2] Agathaeromys praeuniversitatis é conhecido por 35 molares de um único sítio (Seroe Grandi).[3] Embora os depósitos não tenham sido datados com precisão, Zijlstra e colegas sugeriram, com base em correlações com depósitos semelhantes em Curaçau próximo e flutuações do nível do mar, que o material de A. praeuniversitatis provavelmente tem entre 540.000 e 230.000 anos de idade e o de A. donovani provavelmente tem entre 900.000 e 540.000 anos.[22] Sítios fósseis de idade semelhante também são conhecidos das ilhas próximas de Curaçau e Aruba, mas estes contêm vários outros roedores, incluindo Megalomys curazensis e Dushimys larsi em Curaçau e espécies de Oligoryzomys, Sigmodon e Zygodontomys em Aruba.[23] Agathaeromys provavelmente descende de um mmebro de Oryzomyini desconhecido que migrou para a ilha a partir da Venezuela continental em algum momento durante o Pleistoceno.[24]
Referências
- ↑ a b Zijlstra et al., 2010, p. 862; cf. Weksler, 2006; Weksler et al., 2006
- ↑ a b Zijlstra et al., 2010, p. 863
- ↑ a b c Zijlstra et al., 2010, p. 866
- ↑ Hooijer, 1959, p. 18; Zijlstra et al., 2010, p. 861
- ↑ De Buisonjé, 1974, pp. 83, 186–187, 216–217
- ↑ a b c Zijlstra et al., 2010, p. 861
- ↑ Zijlstra et al., 2010, fig. 6
- ↑ Zijlstra et al., 2010, fig. 7
- ↑ Zijlstra et al., 2010, p. 862
- ↑ Zijlstra et al., 2010, pp. 870–871; Turvey et al., 2010, p. 766
- ↑ Turvey et al., 2012, fig. 3
- ↑ Zijlstra, 2012, fig. 3
- ↑ Weksler, 2006, pp. 1, 10; Weksler et al., 2006, p. 1, tabela 1
- ↑ Musser e Carleton, 2005
- ↑ Zijlstra et al., 2010, pp. 862–863
- ↑ Zijlstra et al., 2010, p. 869, tabela 4
- ↑ a b c d Zijlstra et al., 2010, pp. 862, 864, 867
- ↑ Zijlstra et al., 2010, pp. 862, 864–865, 867
- ↑ Zijlstra et al., 2010, pp. 862, 865, 867
- ↑ Zijlstra et al., 2010, pp. 866–867
- ↑ Zijlstra et al., 2010, pp. 867–868
- ↑ Zijlstra et al., 2010, p. 869
- ↑ Zijlstra et al., 2010, p. 872; Zijlstra, 2012, p. 67
- ↑ Zijlstra et al., 2010, p. 872
Bibliografia
- De Buisonjé, P.H. 1974. Neogene and Quaternary geology of Aruba, Curaçao and Bonaire. Uitgaven Natuurwetenschappelijke Studiekring voor Suriname en de Nederlandse Antillen 78:1–291.
- Hooijer, D.A. 1959. Fossil rodents from Curaçao and Bonaire. Studies on the Fauna of Curaçao and Other Caribbean Islands 9(35):1–27.
- Musser, G.G. e Carleton, M.D. 2005. Superfamily Muroidea. Pp. 894–1531 in Wilson, D.E. e Reeder, D.M. (eds.). Mammal Species of the World: a taxonomic and geographic reference. 3rd ed. Baltimore: The Johns Hopkins University Press, 2 vols., 2142 pp. ISBN 978-0-8018-8221-0
- Turvey, S.T., Weksler, M., Morris, E.L., and Nokkert, M. 2010. Taxonomy, phylogeny, and diversity of the extinct Lesser Antillean rice rats (Sigmodontinae: Oryzomyini), with description of a new genus and species. Zoological Journal of the Linnean Society 160:748–772.
- Turvey, S.T., Brace, S. e Weksler, M. 2012. A new species of recently extinct rice rat (Megalomys) from Barbados. Mammalian Biology 77:404–413.
- Weksler, M. 2006. Phylogenetic relationships of oryzomyine rodents (Muroidea: Sigmodontinae): separate and combined analyses of morphological and molecular data. Bulletin of the American Museum of Natural History 296:1–149.
- Weksler, M., Percequillo, A.R. e Voss, R.S. 2006. Ten new genera of oryzomyine rodents (Cricetidae: Sigmodontinae). American Museum Novitates 3537:1–29.
- Zijlstra, J.S., Madern, P.A. e Hoek Ostende, L.W. van den. 2010. New genus and two new species of Pleistocene oryzomyines (Cricetidae: Sigmodontinae) from Bonaire, Netherlands Antilles. Journal of Mammalogy 91(4):860–873.
- Zijlstra, J.S. 2012. A new oryzomyine (Rodentia: Sigmodontinae) from the Quaternary of Curaçao (West Indies). Zootaxa 3534:61–68.
Ligações externas
 Media relacionados com Agathaeromys no Wikimedia Commons
Media relacionados com Agathaeromys no Wikimedia Commons